

12.4.3 Регулирование плотности популяции
Рост популяции регулируется факторами, зависящими и не зависящими от ее плотности.
Не зависящие от плотности факторы, такие как большинство климатических факторов (температура, влажность воздуха, количество осадков и др.), ускоряют или тормозят рост популяции всегда в степени, пропорциональной исходной плотности. Так, например, наступление холодов снизило количество рыбы в трех прудах одинаково на 6-8%, хотя плотность популяций была очень разной (от 55 200 до 2000 рыб на один день ловли) и осталась разной. То же можно сказать и о благоприятных погодных условиях: они могут вызывать массовое размножение независимо от того, высокой или низкой была исходная плотность.
В отличие от этого факторы, зависящие от плотности, как, например, большинство биотических факторов (конкуренция, враги, паразиты, патогенны), влияют на рост популяции только лимитирующим образом, и их действие никогда не бывает одинаковым при разной плотности популяции. Они действуют как регуляторы в узком смысле слова, «подгоняя» популяцию к существующей в данный момент емкости среды. В плотной популяции выше вероятность контактов и распространения инфекций. Отдельный хищник сокращает плотную популяцию жертв меньше, чем редкую, но только в том случае, если он не размножается. Таким образом, действие факторов, зависящих от плотности, сильно зависит также от общей ситуации в экосистеме.
12.5 Экосистемы
Экосистемы – это надорганизменные объединения с определенным видовым составом. В экосистемах суши (лесах, степях, лугах, болотах и т.д.), прежде всего, бросаются в глаза неподвижные, обычно хорошо заметные и обладающие большой биомассой продуценты. Состав растительных сообществ в экосистемах характеризуется, однако, не систематическими категориями, а жизненными формами. Переходы между экосистемами часто постепенны (например, между лесом и степью), границы их нечетки (например, между топяным болотом, тростниковым болотом и озером). Будучи энергетически и структурно открытыми системами, они обязаны своим единством взаимодействию своих компонентов, которые поддерживают в отношении друг к другу и к неживой среде статистическое (подвижное) равновесие. Поэтому экосистемы характеризуются не столько определенными числами и неподвижными границами, сколько средними значениями и границами разброса.
12.5.1 Структура экосистем
Из многих сходных по своей биологии видов одной экосистемы (например, насекомых, сосущих соки луговых растений, – тлей, цикад, клопов) обычно лишь немногие (5-10%) составляют основную часть (около 4/5) биомассы данной группы. Высокая плотность популяций этих доминирующих видов говорит об их оптимальной приспособленности к данной экосистеме и большом для нее значении. Часто они встречаются и в других экосистемах, но образуют там популяции меньшей (субдоминанты) или совершенно незначительной (рецеденты) плотности. В одинаковых экосистемах удаленных друг от друга областей один вид может быть заменен другим: например, в американских тропических лесах среди птиц опылителями цветков служат колибри, а в африканских – медососы. Поэтому экосистему редко можно охарактеризовать перечнем отдельных видов или, в лучшем случае, комбинацией видов. Для этого больше всего пригодны стеноойкные виды, приуроченные к особым условиям данной экосистемы; изредка это бывают доминантные виды (средний дятел в лиственном лесу, клюква на торфяном болоте).
Для вида как структурного элемента экосистемы решающее значение имеет не его систематическое положение (отражающее гомологии в его плане строения), а функциональная конституция и экологическая потенция. Поэтому вид представляет собой комплекс жизненных форм, которые позволяют ему использовать определенные свойства (экологические лицензии) среды (субстраты, источники пищи, диапазоны температур и влажности и т.д.) и играть таким образом определенную роль в экосистеме.
Как один вид может образовывать целый спектр жизненных форм, так каждое местообитание предоставляет организму целый спектр факторов в виде экологических лицензий для использования. Из того и другого – из сочетания области потенции определенной жизненной формы и области валентности необходимых для ее реализации факторов среды – при достаточном совпадении складывается экологическая ниша. Жизненную форму можно сравнить с профессией, имеющуюся валентность – с рабочим местом, а экологическую нишу – с реализованными трудовыми отношениями. Говоря об экологической нише вида, имеют в виду всю многомерную мозаику ниш. И наоборот, каждое местообитание постоянно предоставляет лицензии на ниши для множества организмов. Здесь господствует принцип конкурентного исключения, допускающий в каждую нишу (в узком смысле слова) только один вид.
Возможность внедрения случайно завезенных (колорадский жук) или самопроизвольно распространившихся (кольчатая горлица) видов показывает, что в экосистеме – по историческим и экофаунистическим или экофлористическим причинам – еще могут быть свободные, долго не используемые лицензии. Это может быть связано, например, с тем, что или в область вторжения нового вида еще не проникли противодействующие ему виды, или они были, но исчезли.
Совокупность всех видов, которые встречаются совместно, называют сообществом организмов, или биоценозом, а занятое ими местообитание, т.е. совокупность всех экологических факторов их местонахождения, – биотопом («место жизни»). Но поскольку биоценоз трудно четко отделить от биотопа, лучше объединять их под именем биогеоценоза (рис. 12.13). Биогеоценозу свойственно общее энергетическое и материальное «хозяйство», более или менее отграниченное от других аналогичных комплексов, а также способность к самоподдержанию (путем регуляции).

Рис. 12.13. Биогеоценоз леса (показаны энергетический вход и выход)
Большинство биогеоценозов подразделяется на субъединицы (ярусы, биохоры, меротопы), которые увеличивают возможное число ниш и видов, но в то же время экологически взаимозависимы.
Наиболее стабильны и замкнуты ярусы («этажи»), например, в лесу –древесный ярус, кустарниковый, травянистый и напочвенный. Хотя каждый из них обладает собственным микроклиматом, а часто и собственной фауной (тли, короеды), они связаны между собой климатически и функционально (например, пищевыми цепями с участием листового опада). Многие птицы, гнездящиеся на земле (пеночки и др.), разыскивают себе пищу в кустарниковом и древесном ярусах, а гнездящиеся в кроне (голуби) – на почве.
В менее самостоятельных биохорах (на падали, в экскрементах, на высших грибах, пнях, в гнездах и т.д.) встречаются особые, часто очень богатые видами, но большей частью короткоживущие сообщества, на которые по-разному влияют различные биоценозы, в состав которых входят эти биохоры.
В каждую экосистему входят следующие функциональные компоненты:
а) продуценты – автотрофные организмы, которые с использованием солнечной энергии строят из неорганических соединений богатую энергией биомассу (зеленые растения, сине-зеленые водоросли, фото- и хемосинтезирующие бактерии);
б) консументы – гетеротрофные организмы, которые используют этот органический материал для получения и накопления энергии, изменяют или перестраивают органические вещества (животные и гетеротрофные микроорганизмы);
в) деструенты – гетеротрофы, которые разрушают использованные или отмершие остатки биомассы, разлагают их на неорганические составные части (минерализация), поступающие снова в резервуары минеральных веществ (бактерии, грибы).
Консументы питаются живым (биофаги) или мертвым (сапрофаги) органическим материалом. Среди биофагов могут быть выделены растительноядные организмы, или фитофаги (первичные консументы, к ним относятся также поражающие растения вирусы, бактерии, грибы и паразитические сосудистые растения), хищники (вторичные консументы, в их числе и паразиты первичных консументов) и конечные потребители – вершинные хищники (третичные консументы).
Представители рассмотренных выше трофических категорий (трофических уровней) связаны между собой односторонне направленной передачей биомассы в пищевые цепи (цепи питания). Все пищевые цепи входят в круговороты материи, ведущие от продуцентов к деструентам; цепи могут состоять из малого или большого числа звеньев.
Пастбищные пищевые цепи (рис. 12.14), или цепи хищников, начинаются с продуцентов: клевер – овца – волк; планктонные водоросли – дафния – плотва – щука – скопа. Для таких пищевых цепей характерно увеличение размеров особей (восходящие цепи) при одновременном уменьшении плотности популяций, скорости размножения и продуктивности по биомассе. Цепи паразитов могут начинаться с продуцентов (яблоня – щитовка – наездник) или консументов (овца – муха-кровососка – бактерии – вирусы); для них характерно повышение плотности популяции и скорости размножения при уменьшающихся размерах особей. Детритные цепи проходят через отмершую биомассу; они обычно коротки (листовой опад – дождевые черви – бактерии).
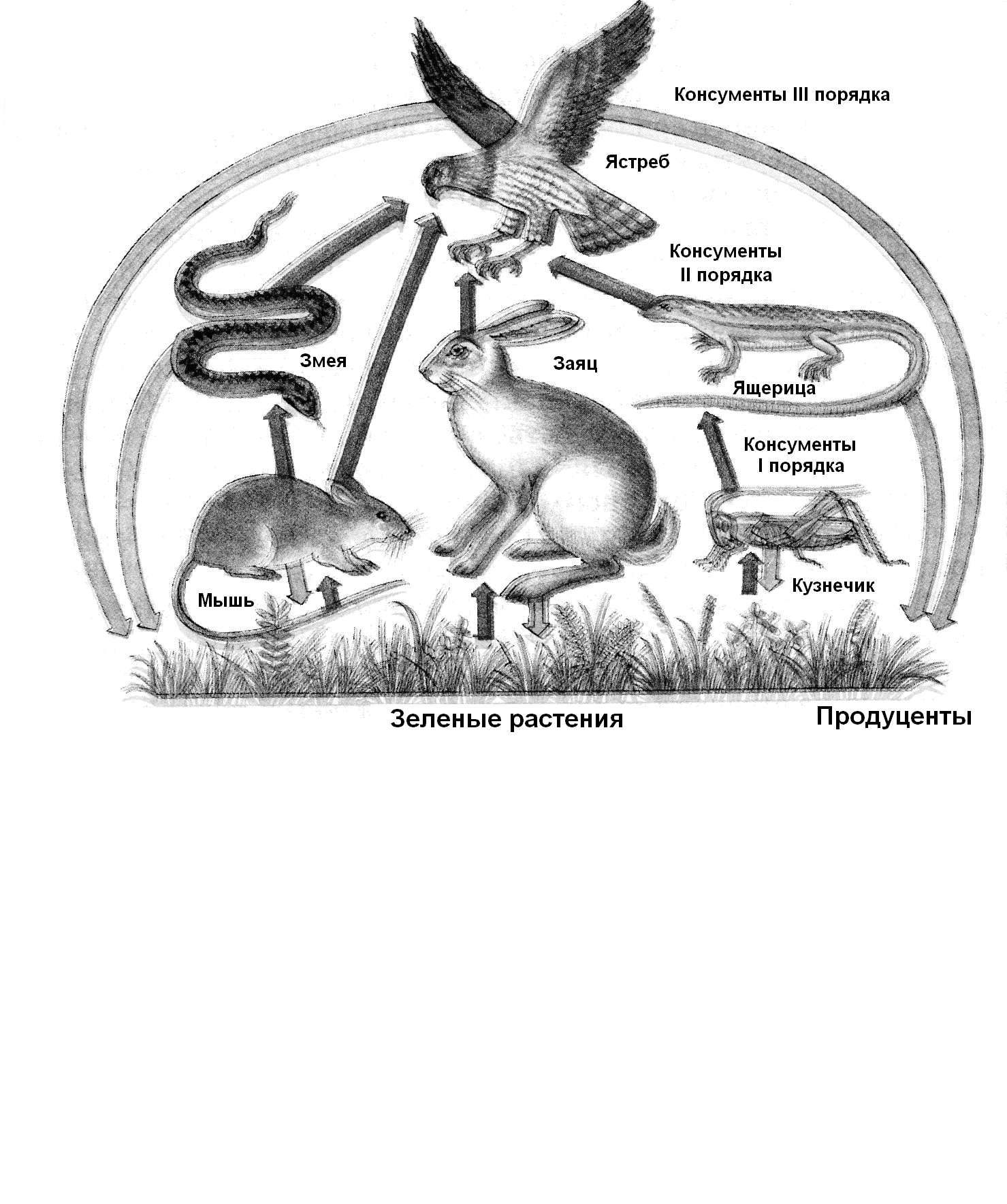
Рис. 12.14. Трофические цепи в наземном биогеоценозе
Так как продуцентов обычно поедают разные консументы, хищники используют много видов жертв, а всеядные организмы (человек, медведь, воробей) потребляют как продуцентов, так и консументов, т.е. живут на разных трофических уровнях, пищевые цепи многократно разветвляются и сплетаются в сложные пищевые сети.
Если расположить друг над другом графические отображения плотности популяций (число особей на 1 м2), биомассы (в граммах сухого или влажного веса на 1 м2) или продуктивности в энергетических эквивалентах (джоулях на 1 м2 в год) для всех членов каждого трофического уровня (в одной пищевой цепи), получится так называемая пищевая (экологическая) пирамида (рис. 12.15). Пирамиды энергии (так как в них учитывается время) стоят всегда «правильно». Напротив, «вверх ногами» стоят пирамиды чисел для цепей паразитов и цепей, начинающихся с древовидных продуцентов.

Рис. 12.15. Экологические пирамиды. А. Кузнечики поедают зеленые растения, лягушки – кузнечиков, змеи - лягушек, орел – змей. Внизу - пирамиды чисел (а), биомассы (б), энергии (в) для упрощенной модельной экосистемы: люцерна, телята, мальчик