

10.2 Рекомбинации
Под рекомбинацией понимают обмен аллелями, перераспределение структур, несущих генетическую информацию, соединение их в новых сочетаниях. Предпосылкой для осуществления рекомбинаций является объединение различного генетического материала. Различают два основных типа такого объединения:
1) половой процесс, при котором происходит слияние генеративных клеток и их ядер;
2) парасексуальный процесс, когда происходит слияние вегетативных клеток, рекомбинация в вегетативных клетках или перенос части генома.
Переносе части генома происходит при:
а) конъюгации у бактерий – контакте между клетками, при котором происходит передача плазмид или частей генома, активированных плазмидами;
б) трансдукции – переносе генетического материала клетки вирусами;
в) трансформации – передаче ДНК через внеклеточную среду;
г) рекомбинации между вирусными геномами.
Объединение различного генетического материала в основном возможно лишь в пределах одного вида или близко родственных видов. Наличие барьеров для скрещивания обусловливает сохранение видов.
У эукариот после объединения различного генетического материала в одном ядре образуются гетерозиготные клетки, по меньшей мере с двумя геномами, различающимися по аллелям одного или нескольких генов. Такие организмы называют помесями или гибридами. У прокариот после переноса генов клетки оказываются диплоидными только по одному гену или какой-то части генома и в случае различия в аллелях называются гетерогенотами.
Гибридное состояние может сохраняться в течение ряда поколений, или же происходит рекомбинация. Рекомбинация может быть
а) на уровне целых хромосом – с образованием нового их сочетания;
б) внутри хромосом – между их частями;
в) между геномом и плазмоном или плазмидами, вирусами и другими дополнительными последовательностями ДНК.
10.2.1 Рекомбинация целых хромосом
Этот процесс определяется двумя фазами:
1) слиянием гаплоидных (в норме) клеток в диплоидную зиготу;
2) координированным распределением хромосом в мейозе.
Эти две ступени определяют характерные типы расщепления после скрещиваний. Впервые эти типы наблюдал Мендель еще в 1865 г., и после их повторного открытия Корренсом, Чермаком: и де Фризом (в 1900 г.) они были положены в основу классической генетики.
При монофакториальном наследовании исходные индивидуумы; (родители – Р) различаются аллелями одного гена. Различные аллели одного гена всегда обозначают одинаковым основным-символом (например, а), чтобы отметить их принадлежность к одному генному локусу. Нормальный аллель, т.е. аллель дикого типа, помечают знаком «плюс» (а+) или А, а мутантный аллель знаком «-» (а–)или а.
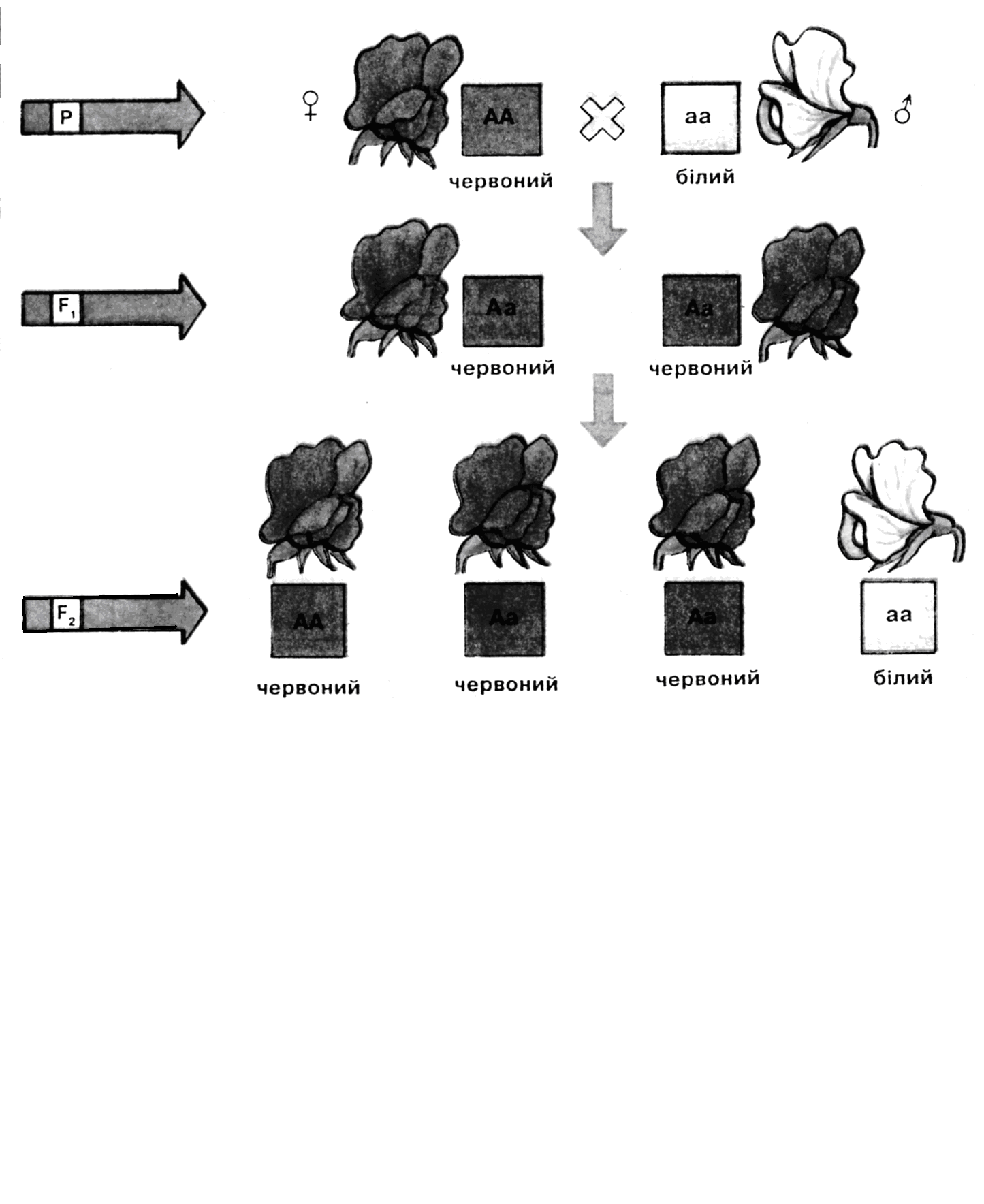
В качестве примера, демонстрирующего результаты моногибридного скрещивания гомозиготных родителей, можно рассмотреть скрещивание растений гороха с различной окраской венчика: красной (доминантной) и белой (рецессивной) Доминантный аллель обозначим А, а мутантный (рецессивный) – а. Цитологические основы расщепления при монофакториальном наследовании представлены на рис. 10.8.
А
Б
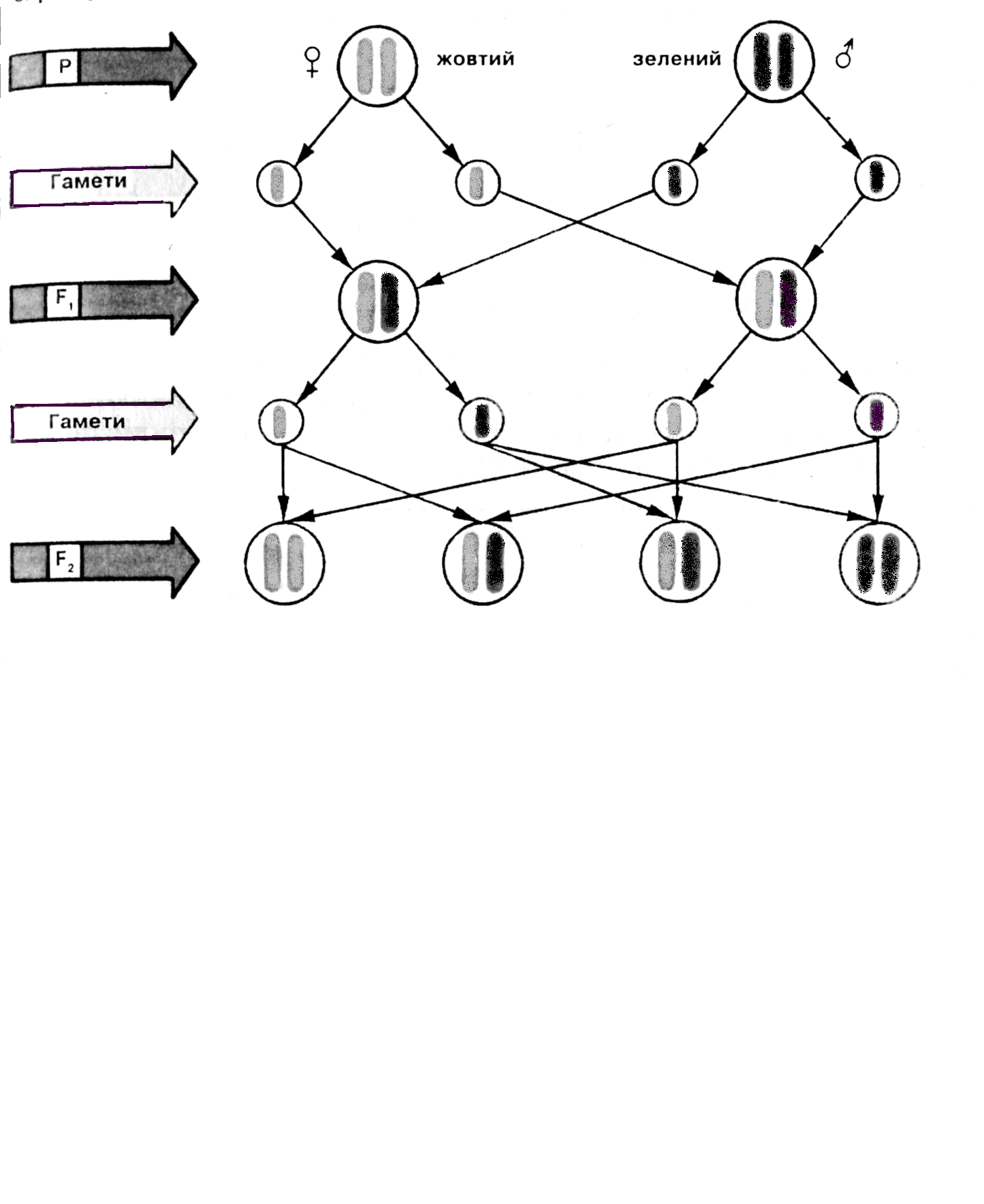
Рис. 10.8. Моногибридное скрещивание. А – результаты скрещивания растений гороха с различной окраской венчика. Б - цитологическое обоснование расщепления при моногибридном скрещивании (хромосомы, которые несут доминантную аллель окрашены в желтый цвет, рецессивную - в зеленый)
Еще одним примером монофакториального наследования с гомозиготными родителями является наследование фенилкетонурии. Нормальный аллель обозначим а+, а мутантный аллель, который обусловливает дефект фермента и тем самым болезнь, — а–. В нашем примере (рис. 10.9) отец гомозиготен с двумя нормальными аллелями, т.е. а+а+, а мать гомозиготна с двумя аномальными аллелями, т. е. а–а–.

Рис. 10.9. Монофакториальное наследование. Пример: рецессивная наследственная болезнь. а+ – нормальный аллель; а– – мутантный аллель, который обусловливает болезнь.
Она здорова только фенотипически благодаря правильной диете. Таким образом, родители различаются по аллелям одного гена (остальные различия мы ради простоты не учитываем). Это относится ко всем соматическим клеткам родителей. При образовании половых клеток диплоидное число хромосом 2n=46 уменьшается до гаплоидного n=23. Аллели вместе с гомологичными хромосомами распределяются так, что все отцовские гаметы содержат аллель а+, все материнские – аллель a-. При оплодотворении ядра гамет сливаются и образуется диплоидная зигота. Аллели а+ и а– объединены теперь в одном ядре, которое становится гетерозиготным (а+а–). Из такой зиготы в результате митозов развивается гетерозиготный эмбрион, и рождается ребенок F1 (первое поколение потомства). Фенотипически ребенок здоров, так как нормальный аллель а+ доминирует над рецессивным мутантным аллелем а-. Так как все комбинации гамет в отношении данного гена должны быть одинаковы, то все дети от этого брака гетерозиготны и здоровы, хотя мать гомозиготна и больна.
В рассмотренном случае больные с наследственным заболеванием имеют с гомозиготным здоровым партнером здоровых детей. В отношении фенилкетонурии сравнительно простым тестом для определения гомо- или гетерозиготности по данному гену служит повышенная нагрузка фенилаланином. Перед вступлением в брак можно с помощью этого теста установить, следует ли учитывать возможность рождения больных детей.
Потомство представителей фенотипически здорового, но гетерозиготного поколения F1 зависит от выбора брачного партнера. На рис. 10.7 представлены три возможных варианта. Когда партнер гетерозиготен (а+а~) или гомозиготен а~а~, могут родиться больные дети; если же он гомозиготен а+а+, то все дети будут здоровыми.
При гетерозиготных родителях (а+а– а+а–) наследование будет таким же, как при скрещивании гибридов F1. У особей F1 все соматические клетки имеют генотип а+ а–. В мейозе с одинаковой частотой образуются гаметы а+ и а–. При образовании зигот гаметы могут с одинаковой вероятностью сливаться в следующих сочетаниях (первой указана мужская гамета, второй – женская): 1) а+ с а+; 2) а+ с а–; 3) а– с а+; 4) а– с а–. Поэтому потомство F2 расщепляется в отношении 1 а+а+ : 2 а+а– : 1 а–а–.
Индивидуумы а–а– будут больны. Вероятность появления больных детей при каждом рождении значительна (1:4), поэтому таким партнерам следовало бы отказаться от возможности иметь детей. Особенно велика опасность появления больных детей при браке гомозиготного больного с гетерозиготным индивидуумом. В этом случае с одинаковой вероятностью можно ожидать рождения больных и здоровых детей (рис. 10.9).
Скрещивания между гетерозиготами (а+а–) и соответствующими гомозиготами (а+а+ или а–а–) называют возвратными: скрещиваниями.
Так, как выше было объяснено на примере фенилкетонурии, у диплоидных растений и животных, включая человека, наследуются все признаки, определяемые одним геном (монофакториально) с доминантным и рецессивным аллелями. При промежуточном наследовании происходит аналогичное расщепление по генотипу, но фенотипы гетерозигот занимают промежуточное положение между фенотипами родителей.
Мендель, основываясь на результатах своих экспериментов по скрещиванию, сформулировал закономерности, известные в настоящее время как «законы Менделя».
Первый закон Менделя (закон единообразия F1): при скрещивании гомозиготных родительских форм в первом поколении потомства все особи однотипны (рис. 10.8, 10.9).
Второй закон Менделя (закон расщепления): после скрещивания потомков F1 двух гомозиготных родителей в поколении F2 происходит закономерное расщепление (рис. 10.8, 10.9). При различии по аллелям одного гена F2 расщепляется по генотипу в отношении 1:2:1. Расщепление по фенотипу зависит от взаимодействия аллелей. В большинстве случаев признак, определяемый аллелем дикого типа, доминирует над мутантными аллелями, реже наоборот. Кроме того, существуют все переходы от доминантности через неполное доминирование до промежуточного соотношения, при котором оба аллеля проявляются в одинаковой мере и фенотип оказывается «усредненным». При монофакториальном наследовании потомство расщепляется по фенотипу в случае полного доминирования в отношении 3:1, а при неполном доминировании – в отношении 1:2:1.
Третий закон Менделя (закон независимого распределения): аллели каждого гена распределяются в потомстве независимо от аллелей других генов.
Этот закон справедлив только для генов, находящихся либо в разных хромосомах, либо в одной хромосоме, но достаточно далеко друг от друга (50 или более единиц карты).
Точного расщепления по Менделю можно ожидать только тогда, когда анализируемое потомство достаточно велико.
Локализацию наследственных задатков в хромосомах и передачу их в соответствии с распределением хромосом впервые удалось продемонстрировать при изучении генетического определения пола у дрозофилы. Определение пола у человека, происходящее аналогичным образом, соответствует схеме возвратного скрещивания (рис. 10.8). F1 расщепляется на 50% ♂ и 50% ♀. У человека реальное соотношение полов при рождении в настоящее время 106 ♂: 100 ♀ (рис.10.10).
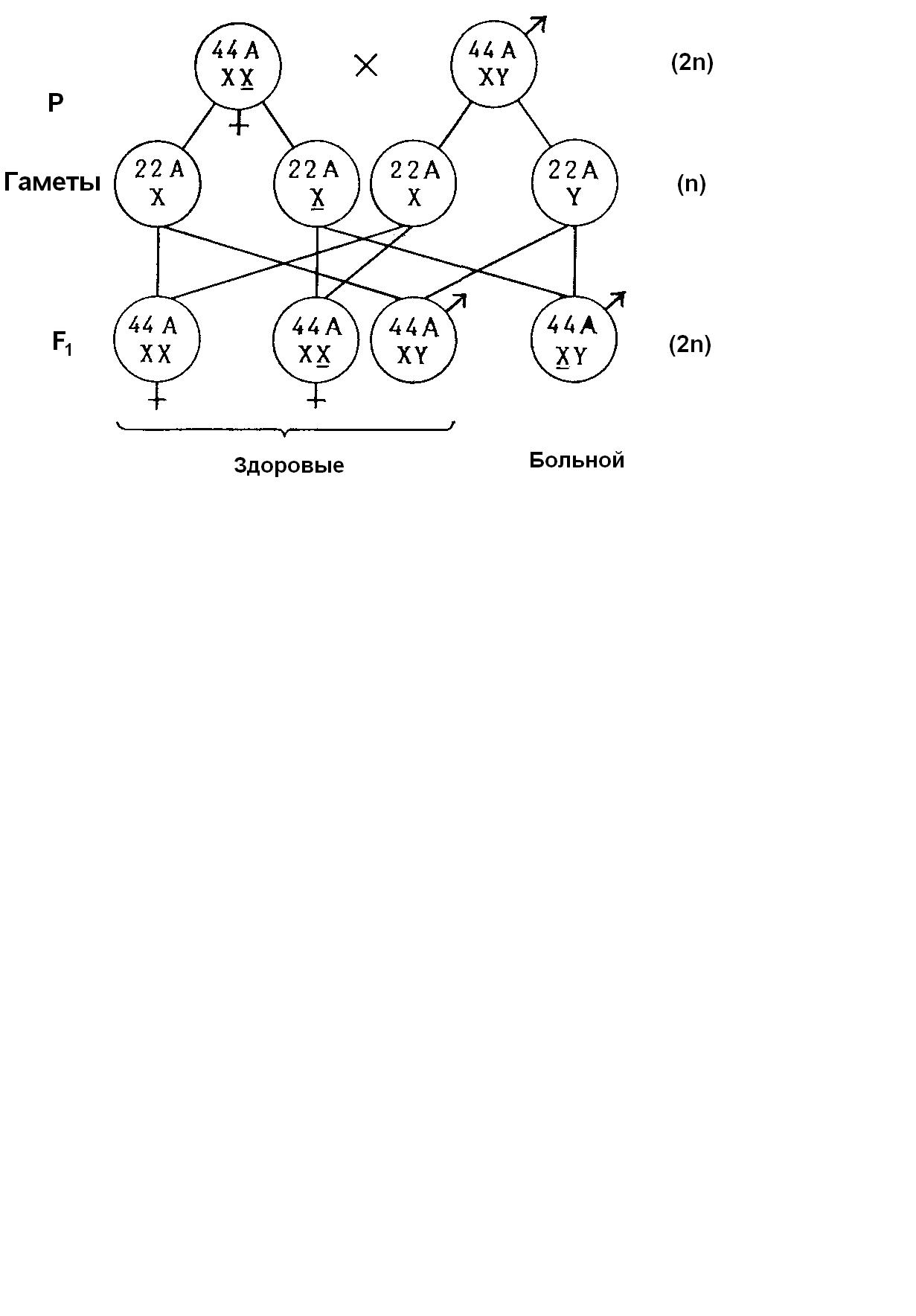
Рис. 10.10. Определение пола и наследование рецессивного заболевания сцепленного с Х-хромосомой, у человека. А – аутосомы, X – Х-хромосома,
Y – Y-хромосома, Х – Х-хромосома с мутантным аллелем болезни крови или дальтонизма
В половых хромосомах локализуются не только гены, необходимые для развития первичных и вторичных половых признаков. В Х-хромосомах находятся, например, гены цветового зрения (дефект: красно-зеленая слепота, или дальтонизм) и свертывания крови (дефект: гемофилия), передающиеся по наследству вместе с Х-хромосомой. В Y-хромосоме соответствующих аллелей нет, и поэтому рецессивные гены, находящиеся в Х-хромосоме, у особей мужского пола проявляются в фенотипе (гемизиготность).