

9.2.5. Мышечное движение
Мышцы состоят из мышечных волокон. Эти волокна могут быть одноядерными или, в случае слияния большого числа мышечных клеток, многоядерными. Мышечное движение основано на способности миофибрилл, лежащих в цитоплазме мышечного волокна, сокращаться в продольном направлении за счет энергии АТР и снова удлиняться.
В так называемой поперечнополосатой мускулатуре, распространенной почти во всем животном царстве от кишечнополостных до позвоночных (скелетная мускулатура, мышцы сердца и языка), в многоядерных мышечных волокнах в световом микроскопе видны попеременные светлые и темные поперечные полосы. Обладающие лишь слабым двойным лучепреломлением (изотропные) светлые диски I разделены пополам поперечной промежуточной линией – пластинкой Z (рис. 9.14). Обладающие сильным двойным лучепреломлением (анизотропные) темные диски А имеют в
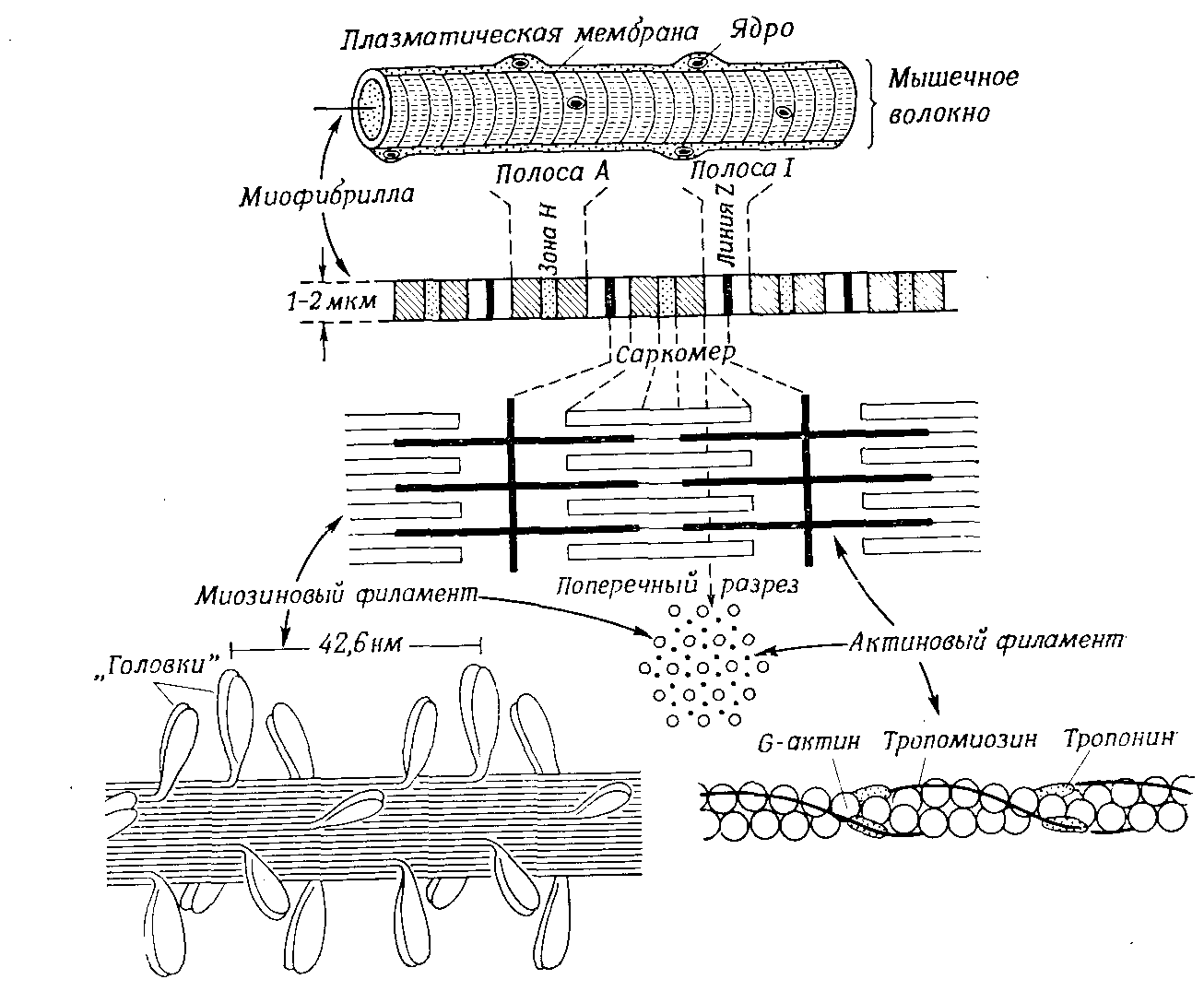
Рис. 9.14. Строение поперечнополосатого мышечного волокна
середине несколько более светлую зону Хензена (зону Н), которая сама может быть разделена пополам срединной линией (линией М). Отрезок мышечного волокна, ограниченный двумя пластинками Z (в мышцах теплокровных его длина около 2 мкм), называется саркомером.
Как показывает электронная микроскопия, миофибриллы состоят из цепей расположенных параллельно элементарных нитей (миофиламентов) двух типов. Более толстые филаменты из L-миозина (диаметр 10 нм) обнаруживаются только в дисках А. Вокруг них группируется по 6 более тонких актиновых филаментов (диаметр 6 нм); они идут от самых пластинок Z и только внутри зон Н заменяются очень тонкими, легко растяжимыми нитями S неизвестного химического состава. Комплекс, образующийся благодаря поперечным связям между филаментами актина и миозина, называется актомиозином.
Мышечные клетки в нормальных условиях активируются импульсами, приходящими по нервным волокнам. В нервно-мышечном синапсе возбуждение передается с нервного волокна на мышечную клетку. При этом каждое мышечное волокно обычно имеет только один нервно-мышечный синапс (двигательную концевую пластинку). Веществом-медиатором, передающим возбуждение, служит ацетилхолин. Он вызывает реакции типа «все или ничего» в виде потенциалов действия, которые активируют сократительный аппарат.
Так же как распространяющееся возбуждение, процесс сокращения подчиняется закону «все или ничего». Это означает, что при надпороговом раздражении амплитуда сокращения волокна всегда одинакова, а если раздражение не достигает порога, она равна нулю. Однако это относится лишь к отдельному волокну, но не ко всей поперечнополосатой скелетной мышце.
Длительное сокращение мышцы в результате суммации одиночных сокращений называют полным тетанусом, если они сливаются на графике в сплошную линию (рис. 9.15). Тетаническое сокращение может быть достигнуто только в том случае, если рефрактерный период достаточно короток.
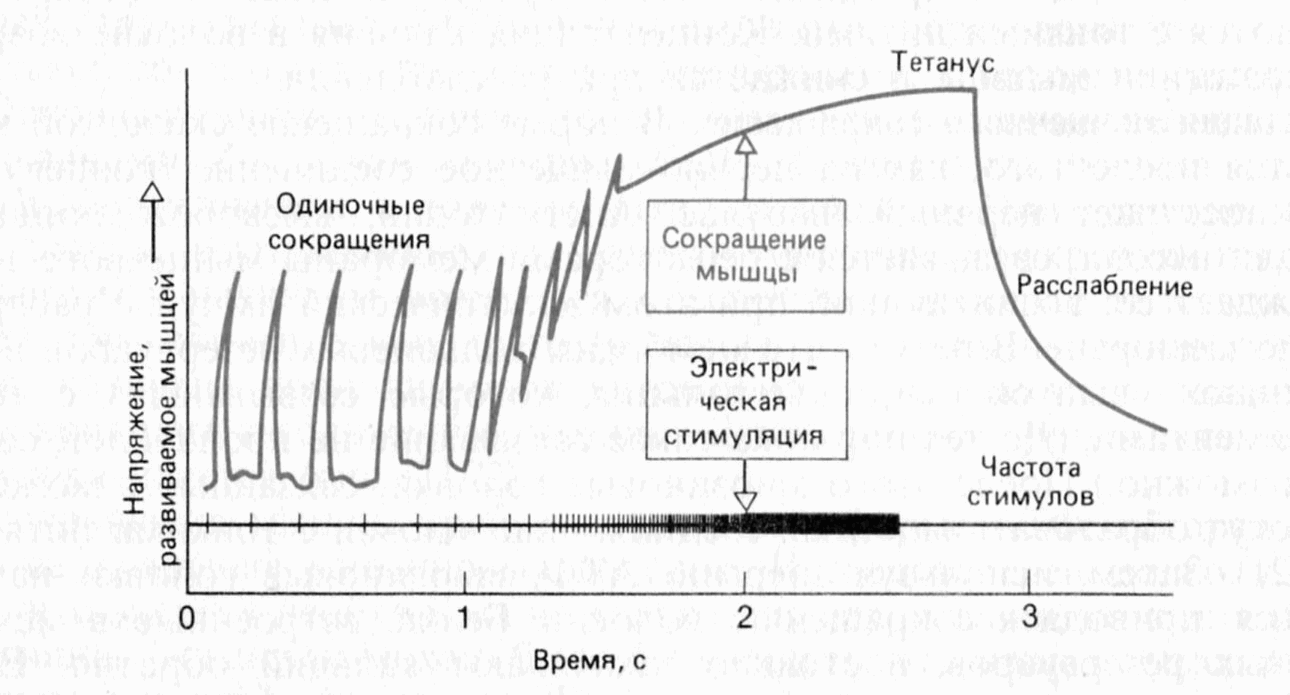
Рис. 9.15. Одиночные сокращения и полный тетанус мышцы
Скелетная мускулатура позвоночных животных обычно все время находится в состоянии некоторого напряжения, поддерживаемого нервными импульсами от двигательных нейронов (тонус). Гладкая мускулатура может долго оставаться в сокращенном состоянии совсем или почти без повышения интенсивности своего метаболизма. В мышечных клетках химическая энергия превращается в механическую работу. Коэффициент полезного действия составляет 30–35%, т.е. около 2/3 энергии теряется в виде тепла.
При сокращение мышечной фибриллы актиновые филаменты вдвигаются между миозиновыми (рис. 9.14, В) и в результате саркомеры укорачиваются. За этот «механизм скольжения» ответственны «головки», имеющиеся на нитях миозина (рис. 9.14). Эти подвижные отростки связываются с определенными местами актиновых филаментов (образуется актомиозиновый комплекс) и сдвигают их в сторону середины саркомера, причем сами отростки наклоняются под углом около 45°. Затем при участии АТР эта связь опять разрывается, отростки быстро выпрямляются, соединяются с другими местами актиновой нити, и процесс повторяется. Этими повторными качаниями головки, подобно веслам шлюпки, подтягивают филаменты актина к середине саркомера. Сложение этих локальных укорочений ведет к сокращению, заметному простым глазом. Без АТР головки застывают на месте, оставаясь прикрепленными к актину под углом 45° (трупное окоченение).