

9.1.5 Научение и память
О научении мы говорим, когда в определенной ситуации, воспринимаемой органами чувств, вероятность определенной формы поведения изменяется в результате предыдущих встреч с той же ситуацией. Некоторые авторы добавляют: изменяется к лучшему, «а именно так, что организм оказывается лучше приспособленным к своей среде» (Эшби). Научение теснейшим образом связано с памятью: в процессе научения происходит перенос информации из внешней среды в хранилище памяти. Способность к научению с несомненностью установлена уже у низших червей (планарий).
Различают следующие типы процессов научения:
1. Привыкание (габитуация). Животные, повторно подвергающиеся воздействию одного и того же раздражителя, за которым не следует никаких биологически значимых событий, реагируют на него все слабее и, в конце концов, совсем перестают реагировать. Так, из-за привыкания прекращается «настораживание» кошки или «замирание» птенцов при определенных звуковых сигналах, вспархивание скворцов с вишневого дерева при выстреле и т.д. Привыкание нельзя объяснить утомлением, оно скорее связано с накоплением торможения, останавливающего реакцию.
Импринтинг (запечатление). Это процесс научения, часто необратимый, протекающий необычайно быстро, в течение более или менее краткого «критического периода». Например, реакция следования у выводковых птиц необратимо закрепляется в первые дни жизни. Фаза запечатления у гусиных птиц заканчивается уже через 12-24 ч после вылупления. У птиц и млекопитающих известно также явление полового импринтинга.
Выработка классических условных рефлексов. Она заключается в связывании «условного» раздражителя (например, слухового или зрительного) с каким-либо рефлексом, ранее с ним не связанным (например, слюнным рефлексом у собаки). Если безусловный раздражитель (например, пища) предъявляют одновременно с условным и такой опыт повторяют многократно, то образуется связь между обоими стимулами и, в конце концов, животное начинает отвечать специфической реакцией (например, выделением слюны) уже на один лишь условный раздражитель, без предъявления безусловного (рис. 9.9).
Выработка инструментальной, или оперантной, условной реакции. Эту форму научения называют также «закреплением действий, ведущих к успеху» или «методом проб и ошибок». Животное может свободно передвигаться в экспериментальных аппаратах (так называемых ящиках Скиннера, лабиринтах, устройствах для опытов с выбором объектов и т.п.) и при этом набирается опыта, усваивая, что определенные спонтанные действия вознаграждаются или позволяют избежать неприятного «наказывающего» раздражителя.
Научение путем подражания. Это сравнительно редкий тип, встречающийся только у высших позвоночных.
Научение в результате «постижения» (инсайта). Решение поставленной задачи с помощью новой комбинации действий приходит здесь внезапно. Этому предшествует фаза относительной неподвижности, во время которой различные возможные действия оцениваются и сравниваются «в уме», без испытания их методом проб и ошибок. Такая форма научения достоверно установлена только у высших обезьян и у человека (рис. 9.10).
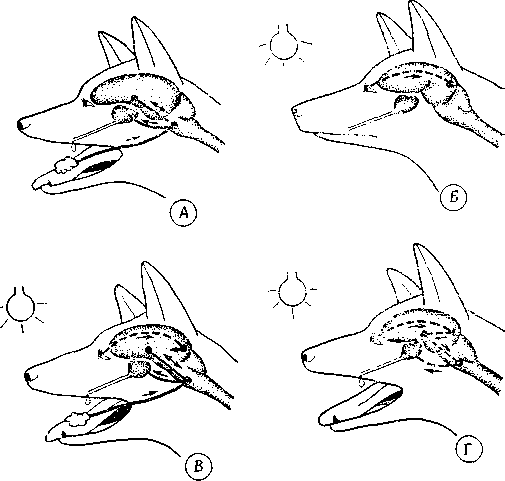
Рис. 9.9. Выработка условного рефлекса. А. Безусловный слюнный рефлекс. Б. Зрительный раздражитель не вызывает реакции. В. Образование условного рефлекса. Г. Условный слюнный рефлекс

Рис. 9.10. Научение типа инсайта
На основании различных наблюдений – главным образом на позвоночных – можно выделить, по меньшей мере, две формы памяти, различающиеся своими механизмами и свойствами. Так называемая кратковременная память у человека имеет максимальную емкость около 160 байт. Длительность сохранения ее следов – от нескольких секунд до нескольких минут. Информация, поступившая в кратковременную память, доступна сразу же после запоминания. Для этой формы памяти характерна чувствительность к помехам. Информация в ней может быть стерта в результате переохлаждения мозга, электрошока, недостаточного снабжения мозга кислородом, наркоза или введения веществ, вызывающих судороги (коразол и др.). Так как во всех этих случаях электрическая активность мозга сильно нарушается или даже совсем прекращается, можно предполагать, что кратковременная память основана на чисто функциональных, электрофизиологических процессах. Вероятно, возникшее возбуждение некоторое время циркулирует по замкнутым нейронным путям, а затем бесследно исчезает, если информация не переносится в долговременное хранилище.
Долговременная память имеет очень большую емкость – от 1010 до 1014 байт. Накопленная в ней информация хранится долго (иногда до конца жизни) и очень устойчива против попыток ее стереть: охлаждение мозга вплоть до исчезновения всякой измеримой активности не приводит к существенной утрате следов памяти. Полагают, что информация в долговременной памяти фиксируется химическим способом. Достоверно показано, что во время обучения в определенных областях мозга увеличивается синтез РНК и, что с процессами научения связаны изменения в синтезе белка. Общее подавление процессов синтеза в нейронах приводит к резко выраженным функциональным нарушениям.
Многие авторы выделяют еще две формы памяти – так называемую промежуточную память, в которой следы сохраняются (у крыс) около 4 часов, и предшествующую кратковременной сенсорную память с еще более коротким временем сохранения следов (несколько сотен миллисекунд).