

11.3.2. Эволюция эукариот
Эта линия, видимо, отделилась от высокоразвитых эобионтов сравнительно рано, параллельно с развитием прокариот. Древнейшие остатки найдены в породах возрастом 1,4 млрд. лет; экстраполяция с учетом скорости замены аминокислот позволяет увеличить оценку возраста эукариот до 2 млрд. лет. Переход к клеткам с хромосомами в отграниченном от цитоплазмы клеточном ядре, с митохондриями и хлоропластами, а также к половому размножению с диплоидией и рекомбинацией расширил эволюционные возможности и стал предпосылкой для многоклеточности и дифференциации.
Относительно происхождения митохондрий и пластид эукариотической клетки существуют две гипотезы: гипотеза эндосимбиоза и гипотеза компартментализации.
Среди одноклеточных эукариот (протистов) есть и авто- и гетеротрофные группы – это следствие раннего расхождения путей развития. Можно думать, что начальным пунктом их эволюции (по крайней мере, если рассматривать водоросли, а возможно, также зоофлагелляты и грибы) были жгутиковые (флагелляты). Для гетеротрофных простейших (Protozoa) исходными могли быть также амебоидные формы; родственные связи здесь неясны, и нельзя исключить полифилию.
Совпадения в составе ассимиляционных пигментов, запасных веществ и в тонком строении хлоропластов указывают на то, что предками высших растений (Cormophyta) были зеленые водоросли. Первыми наземными растениями считаются псилофиты (Psylophyta), жившие 420-350 млн. лет назад (важнейший представитель — Rhynia). Теломная теория пытается вывести особенности строения высших растений, вплоть до семенных, из структуры осевого органа (телома) псилофитов.
Связующим звеном между папоротниками и семенными растениями служат ископаемые семенные папоротники (Pteridospermae). Древнейшие семена найдены в верхнедевонских отложениях. В каменноугольном периоде представлено уже большинство групп растений, но самые высокоразвитые растения (покрытосеменные, Angiospermae) появляются только в конце мелового периода.
Грибы (Fungi) – гетеротрофная, не имеющая хлоропластов группа с сапрофитным, паразитическим или симбиотическим образом жизни – определенно произошли от гетеротрофных примитивных эукариот, стоявших близко к истокам главной линии царства животных. Грибы часто рассматривают как третью крупную группу многоклеточных наряду с Metaphyta и Metazoa.
Животные – вторичные гетеротрофы и облигатные аэробы; как первоисточник пищи им необходимы автотрофные организмы, а также требуется достаточное снабжение кислородом (точка Пастера 0,2% О2).
Древнейшие находки многоклеточных животных – автралийская фауна Эдиакары (возраст 600-700 млн. лет), время которой совпадает с достижением точки Пастера. Это кишечнополостные, кольчатые черви и членистоногие, почти все без твердых образований. Важные опорные пункты для гипотез о происхождении многоклеточных животных дает сравнительная эмбриология. Несомненно, все эти животные (может быть, за исключением губок) развились из одного корня (монофилетически). Согласно разным теориям, исходной группой могли бы быть амебы, жгутиковые или инфузории (рис. 11.15). Теория гастреи исходит из объединения отдельных жгутиковых в колонию, которая преобразуется сначала в бластулоподобный полый шарик (по другой версии – в уплощенную плакулу), а затем путем впячивания – в двуслойное образование с первичной кишкой – гипотетическую гастрею. Из гастреи можно вывести губок (Porifera) и стрекающих кишечнополостных (Cnidaria), а из последних – всех остальных многоклеточных животных.
А |
|
Б |
|
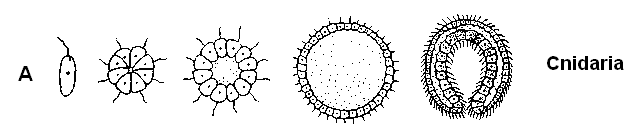
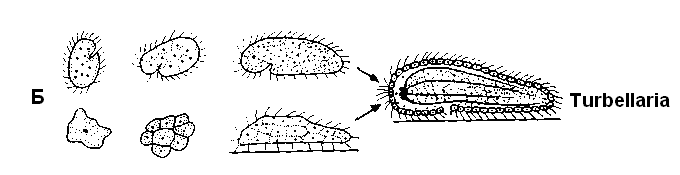
Рис. 11.15. Различные представления о происхождении многоклеточных животных (схемы). А. Теория гастреи: колония жгутиконосцев – двуслойная гастрея. Б. Ацельная теория: от предков-инфузорий к бескишечным турбелляриям (Acoela). В другом варианте (нижний ряд) за исходные формы принимаются амёбы (По Reisinger с изменениями)
Для всех животных, стоящих выше кишечнополостных (стрекающих и гребневиков), т.е. для всех Coelomata (Bilateria), характерны наличие третьего зародышевого листка, вторичная полость тела (целом, иногда редуцированный), все большая способность к активному передвижению, прогрессивная дифференциация нервно-сенсорного аппарата, возрастающая роль переднего полюса тела как конца, направленного при движении вперед, с ротовым отверстием и органами чувств.
Среди современных Coelomata исходной группой считаются архицеломаты. От них идут две основные линии с более развитым членением тела (метамерией): Gastroneuralia (животные с брюшной нервной цепочкой) и Notoneuralia (животные со спинной нервной цепочкой). Среди Gastroneuralia особенно высокого развития достигли на суше представители членистоногих – насекомые, а в море представители моллюсков – головоногие (Cephalopoda). Notoneuralia достигли своей вершины в позвоночных (Vertebrata) (рис. 11.16). В обеих крупных ветвях успешно завершен выход на сушу (многие группы членистоногих, в какой-то степени моллюски, четвероногие позвоночные – Tetrapoda).
Периоды
|
|
Антропогеновый |
|
Неогеновый |
|
Палеогеновый |
|
Меловой |
|
Юрский |
|
Триасовый |
|
Пермский |
|
Каменноугольный |
|
Девонский |
|
Силурийский |
|
Ордовикский |
|
Кембрийский |

Рис. 11.16. Дендрограмма позвоночных
Таблица 11.2