

12.5.2 Физиология экосистем
Экосистема - открытая система, в которой процессы образования и дальнейшей передачи биомассы идут за счет притока энергии солнечного излучения. При каждой передаче на следующий трофический уровень часть доступной энергии не воспринимается, часть отдается (в виде тепла), а часть расходуется на дыхание. По грубой оценке, при этом общая энергия каждый раз уменьшается в десять раз, а при передаче от первичных продуцентов консументам – даже в сто раз (рис. 12.4). Следствие этого – ограниченная длина пищевых цепей, а также уменьшение количества особей и общей энергии при переходе от низших уровней к высшим. Чем длиннее пищевая цепь, тем меньше остается к ее концу доступной энергии.
12.5.3 Развитие экосистем
Хотя экосистемы, находясь в стационарном состоянии, отличаются статистическим постоянством своих элементов, они могут развиваться, переходя от более простых к более сложным формам.
Там, где отступающее море, высыхающее озеро, ледник или деятельность человека освобождают девственную сушу, а также там, где из вулканического пепла, из лавы или в результате выветривания горных пород образуется новая почва, начинается ее заселение редкой, одноярусной пионерной растительностью. С накоплением гумуса и пыли, с повышением влажности почвы развиваются двухъярусные вторичные сообщества, луга и степи. Устойчивая конечная стадия (климакс) этой первичной сукцессии – в большинстве случаев многоярусный лес (рис. 12.16). Вторичные сукцессии как бы залечивают повреждения, естественные или нанесенные человеком (последствия бури, вырубки, пожара, наводнения, выпаса скота). Каким будет конечный этап сукцессии, зависит прежде всего от макроклимата (климатический климакс). Например, на больших высотах и в высоких широтах климаксным сообществом будет уже не лес, а тундра. На сукцессию влияют также эдафические (почвенные, водные) и топографические особенности (уклон, экспозиция, уровень грунтовых вод). Так же может влиять деятельность человека – выпас скота, сенокос, полеводство (параклимакс), так что ландшафты чаще всего представляют собой мозаику из экосистем с разной степенью зрелости.
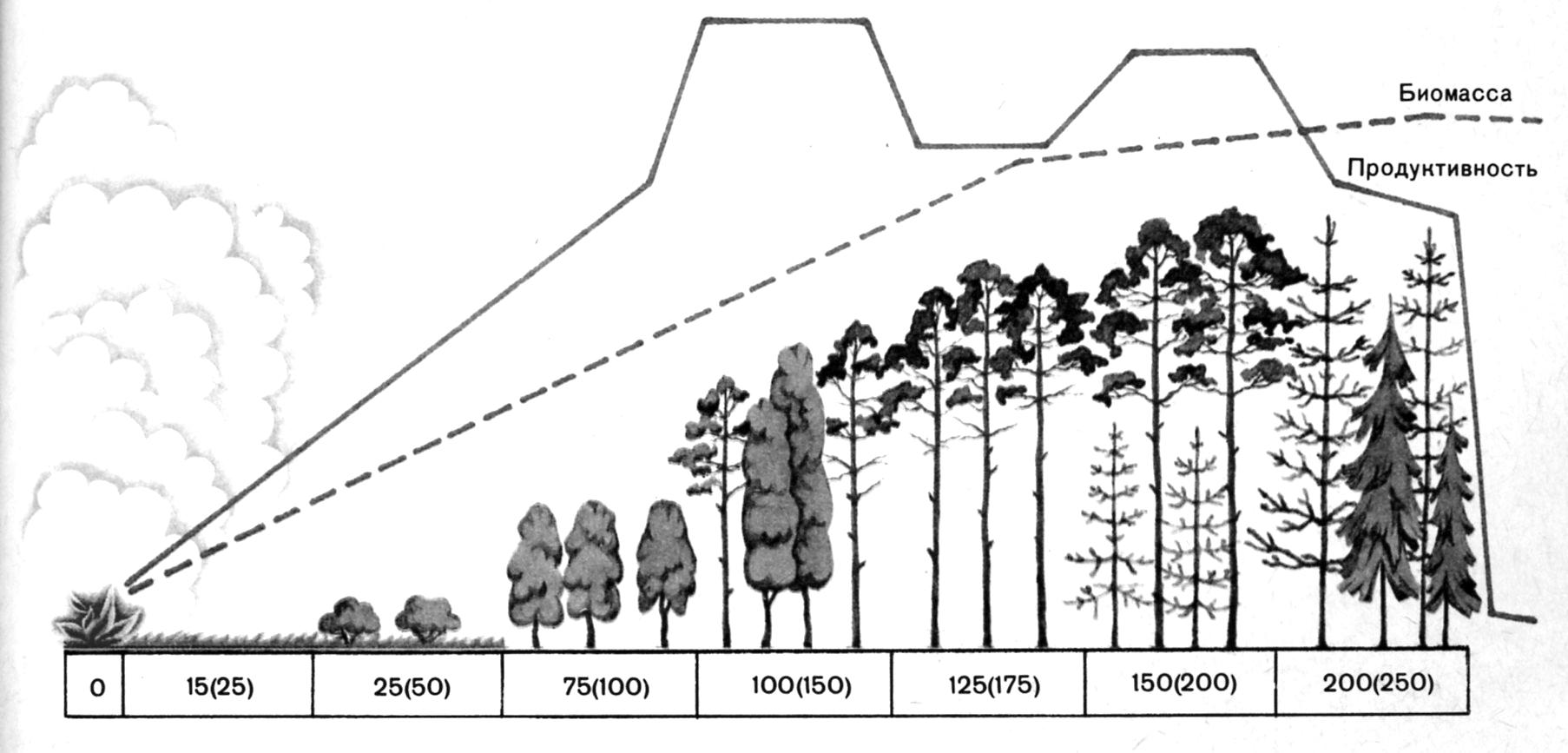
Рис 12.16. Схема сукцессии
В молодом, преимущественно автотрофном лесу производится излишек биомассы (Рп), который может накапливаться в виде древесины. Создающиеся ниши заселяются гетеротрофами, которые все больше используют прирост биомассы. Наконец, в климаксном лесу складывается сложная пищевая сеть с возрастающим удельным весом детритных пищевых цепей, в которых используется вся продукция. Поэтому человеку удается снимать высокие урожаи биомассы только на ранних стадиях сукцессии, когда нетто-продукция велика. В земледелии и лесном хозяйстве используются начальные фазы экосистем с немногими (предпочтительно одним) первичными продуцентами (монокультуры). Конечно, за это приходится расплачиваться неустойчивостью пионерных сообществ по отношению к внешним факторам и конкурирующим консументам. С гетеротрофами, желающими найти себе нишу в такой экосистеме, приходится бороться как с «вредителями».
Сукцессии экосистем, как и эволюция живого на всех уровнях, направлены на обеспечение дальнейшего существования, на гомеостаз. Очевидно, что стабильность подвижного равновесия при меняющихся условиях среды легче всего достигается в том случае, если экосистема состоит из максимально возможного числа компонентов (внутреннее разнообразие); тогда экологические потенции разных видов могут так дополнять друг друга, что разные помехи, как внешние (особенно непредсказуемые изменения абиотических факторов, к которым невозможно приспособиться), так и внутренние (чрезмерные репродуктивные потенциалы некоторых организмов), будут сглаживаться.