

12.3.2 Водный режим
Обменные процессы - биохимические реакции, которые протекают в водных растворах. Поэтому для активной жизни необходимо достаточное содержание воды в организме (у наземных животных 45-95% веса тела). Так как давление водяного пара в воздухе, как правило, сравнительно невелико, неизбежны потери воды в результате транспирации, а покрыть все тело водонепроницаемой изоляцией (кутикулой, слоем воска, роговым панцирем) невозможно из-за необходимости газообмена при дыхании и фотосинтезе. Поэтому запас воды приходится часто пополнять путем питья (позвоночные, пауки, брюхоногие моллюски) или потребления влажной пищи (виды, питающиеся листьями и плодами). Обитатели аридных районов используют метаболическую воду, образующуюся при окислении жиров и других веществ; жир запасается, например, в горбах верблюдов. При высокой влажности воздуха возможно поглощение влаги через покровы тела (амфибии, брюхоногие моллюски, тропические растения-эпифиты), иногда за счет гигроскопичности каких-либо структур – против градиента влажности (мокрицы, клещи, мучной хрущак). Растения аридных местообитаний (пустынь) обладают очень широко расходящимися или очень глубоко проникающими в землю корнями с низким (сильно отрицательным) осмотическим потенциалом.
Потери на транспирацию уменьшаются благодаря защитным покровам и подушкам неподвижного воздуха (удерживаемого шерстью и перьями у животных, волосками у растений), обратному всасыванию воды при выделении мочи и дефекации, поискам влажных мест. Муравьи, термиты, комары активно разыскивают подходящие слои воздуха (гигротаксис). Большинству видов свойственно фоботаксическое (гигрокинетическое) поведение: их ненаправленная активность успокаивается только в месте с подходящей влажностью воздуха. Гигрофильные виды почти не имеют защиты от высыхания и могут существовать только в очень влажном воздухе (амфибии, безраковинные брюхоногие моллюски, большинство обитателей почвы, травы тропического дождевого леса). Ксерофильные виды обладают либо морфологическими средствами защиты от высыхания, либо могут накапливать воду (пустынные суккуленты, например кактусы), всасывать ее обратно, в сухое время суток разыскивать укрытия или восполнять потери воды большим количеством влажной пищи (рис. 12.8).

Рис. 12.8. Растения, принадлежащие к различным экологическим группам
12.3.3 Фактор света
Являясь источником энергии для фотосинтеза, свет имеет фундаментальное экологическое значение. Даже более или менее лишенные света экосистемы глубинных морских вод, почв и пещер в конечном счете зависят от первичной продукции биомассы зеленых растений на свету.
L – освещенность в данном местообитании по сравнению с полной освещенностью непокрытой почвы. Светолюбивые растения пустынь, степей, тундр и высокогорья могут ассимилировать только при L = 100%. У теневыносливых растений точка компенсации (при которой ассимиляция СО2 равна высвобождению СО2) достигается при гораздо меньшем световом довольствии, а неослабленный свет им вреден (рис. 12.9). Луговые растения занимают среднее положение. На состав растительного покрова сильно влияют и видоспецифические требования растений к освещенности, и местные условия освещения. Лесной бук, теневыносливый и в молодом возрасте, вытесняет светолюбивые дубы и сосны; под пологом его крон могут существовать только теневые растения.
Свет вызывает в органах растений (стеблях и их разветвлениях, черешках листьев и т.д.) движения типа искривлений, направленные на оптимальное использование света листьями, т.е. наивысшую активность фотосинтеза (фототропизм).
Так называемым растениям длинного дня (в основном это растения средних широт или происходящие оттуда) для начала цветения требуется, в общем, более длительный светлый период суток (>12-14 ч в сутки), чем растениям короткого дня, происходящим из экваториальных областей (<12 ч в сутки), поэтому последние могут у нас цвести только осенью (или весной).
12.3.4 Экологическая потенция
По экологической потенции в отношении температуры, света, влажности воздуха, содержания солей и кислорода в воде и т.д. различают виды с широкой и узкой нормой реакции: эвритермные и стенотермные, эвригидрические и стеногидрические эвригалинные и стеногалинные, эвриоксибионтные и стено-оксибионтные. Лишь у немногих видов требования ко всем или большинству факторов среды так малы, а широта реакции соответственно так велика, что они могут существовать практически во всех местообитаниях данной области, то есть быть эвриойкными. Полностью эвриойкные виды и настоящие космополиты редки (человек, крыса). Стеноойкные виды специализированы для жизни в определенных местообитаниях; но эврипотентные виды тоже могут оказаться стеноойкными в тех областях, где их требования, например к температуре, удовлетворяются только в отдельных специфических местообитаниях.
В центре ареала данного вида диапазон его потенции часто в значительной мере совпадает с широтой валентности большинства факторов среды, а на окраинах ареала, напротив, тот или иной фактор часто приходится на область пессимума.
Редко бывает так, чтобы в одном месте области валентности всех жизненно важных факторов совпадали с диапазонами потенций вида. Чаще всего хотя бы один фактор лежит вне оптимума. Тогда от этого фактора зависит возможность существования вида в данном месте. Экологически решающим всегда оказывается фактор, оказавшийся пессимальным для той стадии, которая обладает наименьшей широтой реакции.
Выход за пределы пессимума не всегда опасен для индивидуума. Возможны два способа «эмансипации» от экологической валентности: сдвиг диапазона потенции путем акклиматизации (например, спячка) и уклонение от неблагоприятного фактора среды посредством миграции.
Успешность акклиматизации к низким температурам (резистентность к холоду) у пойкилотермных (холоднокровных) животных зависит от предшествующего опыта. У гомойотермных животных снижение температуры действует не только закаливающим образом, но и способствует образованию подкожных жировых отложений и более густого меха.
У холоднокровных и примитивных теплокровных животных пессимальные условия среды вызывают спячку, т.е. прекращение активности и замедление процессов метаболизма и развития, немедленно и неизбежно. Гораздо чаще спячка наступает лишь после более длительного воздействия неблагоприятных условий – олигопаузы, часто на вполне определенной стадии (на стадии яйца, личинки, куколки, у взрослого животного) (рис. 12.9). При этом начало и окончание паузы часто определяются уже не одним только пессимальным фактором, а в основном длиной светового дня (фотопериодом). Длина дня, регулярно изменяющаяся на протяжении года, предвещает приближение благоприятных и неблагоприятных сезонов точнее, чем все другие, менее регулярные колебания климатических факторов.
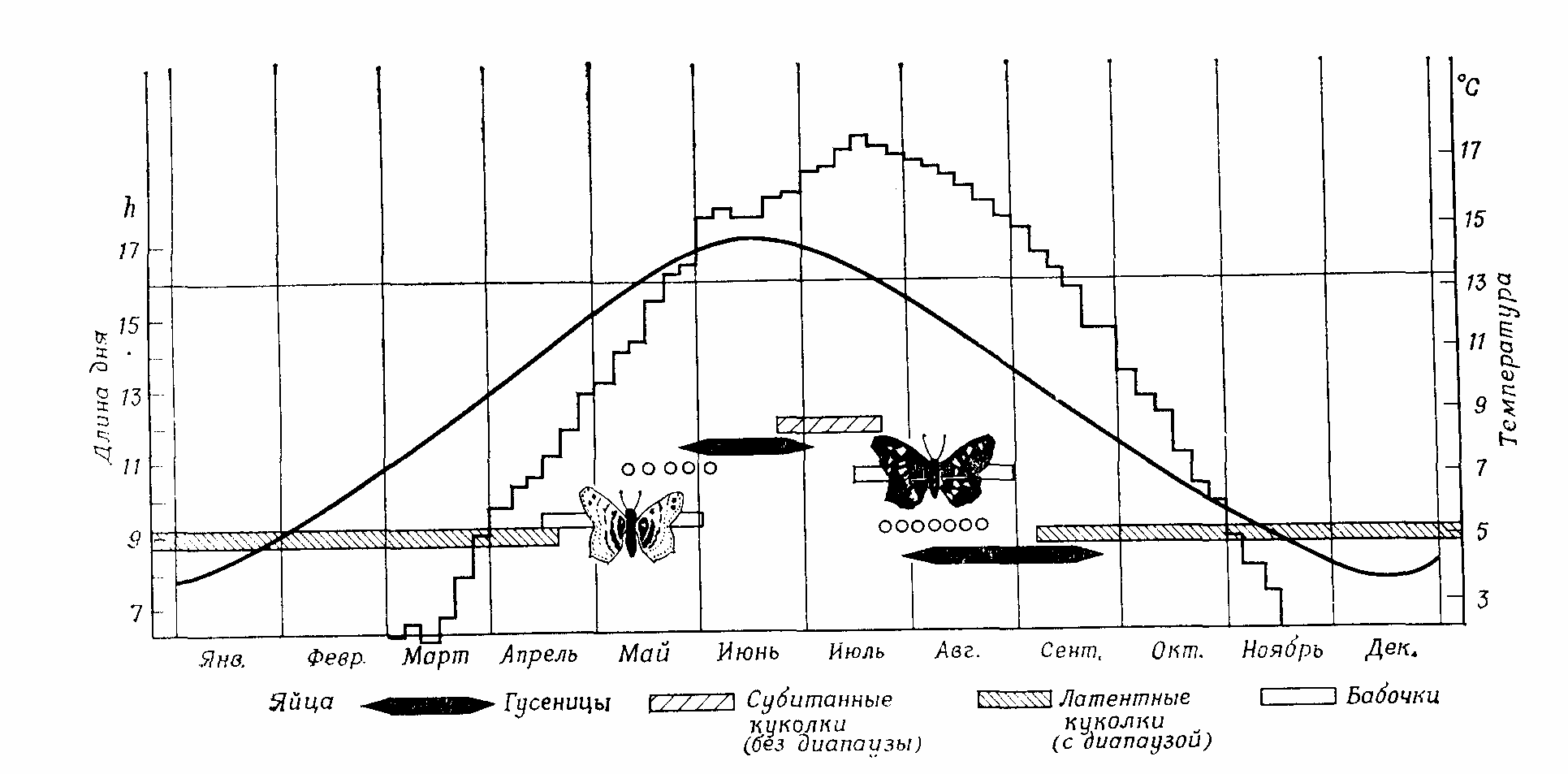
Рис. 12.9. Фотопериодическая индукция диапаузы: возникновение сезонных форм Araschnia levana в результате развития куколок с диапаузой или без нее. Вариант развития определяется длиной светового дня на стадии гусеницы. Ступенчатая линия – средние температуры за пятидневку. Плавная кривая – длина дня (включая «гражданские сумерки»). Горизонтальная линия – критический фотопериод. (По Miiller, с изменениями.)
Ухудшение условий среды может также приводить к миграциям, причем последние иногда совершаются заблаговременно. Недостаток пищи или ухудшение погоды побуждает некоторых летающих насекомых (саранчу), птиц (клеста, кедровку, свиристель), а также млекопитающих (леммингов) к первоначально ненаправленным откочевкам. Регулярные миграции многих перелетных птиц определяются ежегодно изменениями погоды (так называемые погодные птицы – грачи, лысухи, зяблики, дрозды и др.). Другие («инстинктивные») птицы отлетают в более или менее фиксированные сроки, руководствуясь, по-видимому, фотопериодом, задолго до ухудшения погоды и сокращения пищевых ресурсов (таковы, например, кукушка, черные стрижи, аисты, иволга).