

Изменение аллеля дикого типа и его продуктов (mPhk и полипептидной цепи) в результате вставки и делеции
Структуры |
Часть последовательности |
|
Аллель дикого типа |
ДНК
mPHK |
-C-C-C-G-G-T-A-G-C-C-C-C- -G-G-G-C-C-A-T-C-G-G-G-G-
-C-C-C-G-G-U-A-G-C-C-C-C- |
Полипептидная цепь |
— Pro — Gly —Ser —Prо — |
|
После вставки |
ДНК
|
+С -C-C-C-C-G-G-T-A-G-C-C-C-C- -G-G-G-G-C-C-A-T-C-G-G-G-G-
-C-C-C-C-G-G-U-A-G-C-C-C-C- |
mPHK |
||
Полипептидная цепь |
— Pro — Arg — amber (конец цепи) |
|
После делеции |
ДНК
|
-G -C-C-C-G-T-A-G-C-C-C-C- -G-G-G-C-A-T-C-G-G-G-G- -C-C-C-G-U-A-G-C-C-C-C- |
mPHK |
||
Полипептидная цепь |
— Pro —Val — Ala —Pro |
|
Изменение может привести либо к снижению или повышению активности, либо к полной ее потере, но может и не оказывать влияния на признак (в последнем случае мутацию большей частью не удается выявить). Как изменяется признак, зависит от места мутации в гене и от структуры измененной полипептидной цепи
Большинство генных мутаций дают отрицательный эффект и обусловливают выпадение какой-либо ферментативной активности наследственным заболеванием, могут сильно различаться по выраженности симптомов болезни.
У диплоидов мутации возникают только в одном из двух аллелей. В результате получаются гетерозиготы, у которых фенотип определяется взаимодействием аллелей.
Накопление вредных продуктов из-за генетической блокады того или иного пути метаболизма служит причиной многих наследственных болезней человека, например фенилкетонурии (накопление аминокислоты фенилаланина). Это одна из немногих наследственных болезней, при которых фенотипические проявления (слабоумие) можно существенно уменьшить. Так как фенилаланин в нашем организме не синтезируется его накопление можно предотвратить, соблюдая диету с минимальным содержанием фенилаланина. В результате достигается фенотипическое излечение, при котором, однако, сам ген не изменяется; мутантный аллель от таких больных передается по наследству.
Индуцировать генные мутации могут ультрафиолетовые лучи, ионизирующие излучения и химические мутагены.
Из химических мутагенов чаще всего применяют алкилирующие агенты. К ним относятся: этилметансульфонат, нитрозогуанидиновые соединения, аналоги азотистых оснований, такие как бромурацил и 2-аминопурин, а также азотистая кислота и гидроксиламин. Мутации со сдвигом рамки индуцируются акридином.
Только в немногих случаях мутации реализуются сразу же после воздействия физического или химического мутагена. Часть повреждений устраняется в результате репаративных процессов. При безошибочной репарации вновь восстанавливается нормальное исходное состояние, тогда как ошибка при репарации может, например, привести к замене основания. Можно предположить следующие пути возникновения генных мутаций:
а) одно основание превращается в другое (замена оснований);
б) ДНК изменяется так, что начинается процесс ее репарации, при котором включается «не то» основание;
в) несоответствующий нуклеотид включается в результате ошибки репликации.
Все мутагены вызывают повреждения нескольких типов. Так, при действии азотистой кислоты (АК) сначала цитозин дезаминируется в урацил, в результате пара C-G заменяется на Т-А. Кроме этого, азотистая кислота вызывает в ДНК повреждения, которые только после репарации становятся мутациями.
![]()
![]()
![]()
![]()
![]()
![]()
![]()
Мутации возникают с частотой 10-5-10-10 (на один ген). С такой же частотой некоторые из мутантов могут снова превращаться в дикий тип. Либо возникшее изменение устраняется в результате истинной обратной мутации, либо признак дикого типа восстанавливается вследствие второй, так называемой супрессорной, мутаций.
Возникновение мутаций при репликации зависит от частоты ошибок при включении комплементарных нуклеотидов ДНК-полимеразой. Такие мутации встречаются очень редко.
В интактных клетках существуют различные «ремонтные» системы, устраняющие повреждения, вызванные облучением или химическими мутагенами. В репаративных процессах различают фотореактивацию, эксцизионную репарацию и пострепликативную репарацию.
Лучше всего изучена репарация повреждений, вызванных ультрафиолетовыми лучами (репарация других повреждений частично происходит аналогичным образом). При облучении ультрафиолетом между соседними пиримидиновыми основаниями одной цепи возникают димеры, чаще всего димер Т-Т, т.е. вместо водородных связей между Т и А двух нуклеотидных цепей образуются связи Т-Т внутри одной цепи (рис. 10.7).
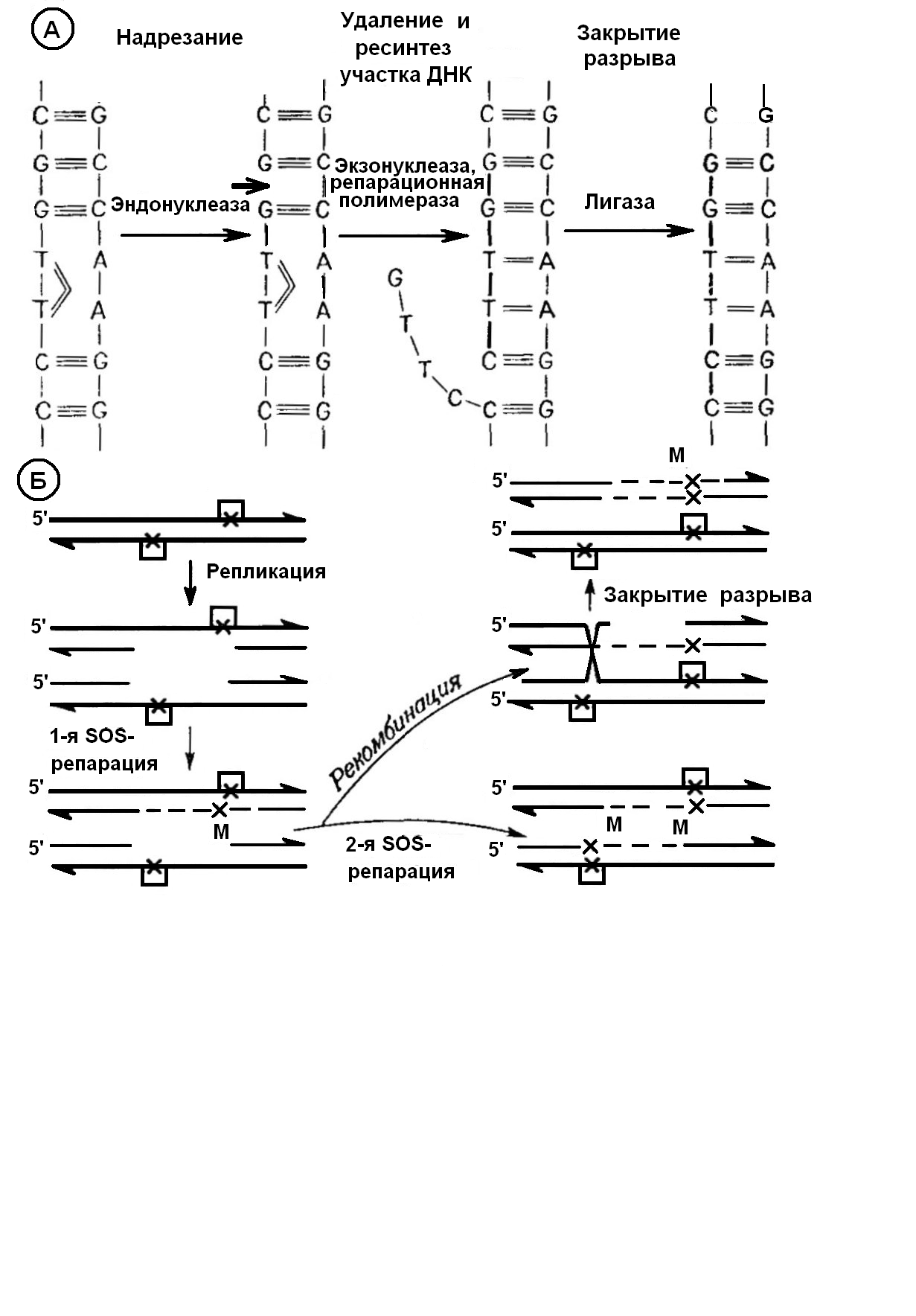
Рис 10.7. Репаративные процессы. А. Эксцизионная репарация (на примере Escherichia coli). Б. Пострепликативная репарация. В представленном примере разрыв в одной молекуле ДНК закрывается путем SOS-репарации, причем возникает мутация (М). Во второй молекуле ДНК разрыв может быть тоже заполнен путем SOS-репарации или закрыт путем рекомбинации с последующим репаративным синтезом, при котором матрицей служит интактная цепь ДНК. (По Bohme, Adler, с изменениями)
Фотореактивация происходит при воздействии видимого света. При этом репаративный фермент разделяет димер на мономеры и опять восстанавливает водородные связи Т-А между комплементарными цепями.
Эксцизионная и пострепликативная репарация не зависит от света, и поэтому ее называют также темновой репарацией.
При эксцизионной репарации (лат. excisio – вырезание; рис. 10.7) вырезается поврежденный участок ДНК. Сначала эндонуклеаза разрезает одну цепь. Следующий фермент – экзонуклеаза – удаляет измененную часть, а ДНК-полимераза вновь синтезирует в направлении 5'→3' недостающий участок, комплиментарный к антипараллельной цепи ДНК. И, наконец, свободные концы старой части цепи соединяются с концами вновь синтезированного участка при помощи лигазы. Фотореактивация и описанная здесь эксцизионная репарация коротких участков ДНК протекают без ошибок и, следовательно, не ведут к мутациям.
Если димеры не будут устранены, то соответствующие основания не смогут выполнять роль матрицы, и в этих местах во вновь синтезированной цепи ДНК окажутся пропуски. Путем рекомбинации между двумя двойными цепями ДНК – продуктами репликации – возможно образование одной нормальной двойной цепи (пострепликативная репарация). Когда после репликации в результате такой рекомбинации получается новая интактная двойная спираль ДНК, процесс репарации тоже не ведет к появлению ошибок. Если повреждения лежат так тесно друг возле друга, что пропуски перекрываются, тогда для заполнения пропусков используется другая «ремонтная» система (SOS-репарация), способная синтезировать новую цепь ДНК и на дефектной матрице. При этом новые основания включаются таким образом, что могут возникнуть мутации (рис. 10.7).
Большинство мутантов уступают дикому типу в приспособленности, менее жизнеспособны и поэтому отсеиваются в процессе отбора. Для селекции и эволюции большое значение имеют сравнительно редкие мутанты с благоприятными или нейтральными изменениями. Чтобы иметь достаточное число мутантов, можно с помощью мутагенов повышать частоту мутаций до 10-2.
Человек, напротив, должен оберегать себя от неблагоприятных изменений, к которым могут приводить мутации. Каждая мутация, затрагивающая ткань, из которой образуются половые клетки, будет передаваться последующим поколениям.