

12.4 Популяция и окружающая среда
Решающие биотические элементы естественных экосистем – это не особи, а популяции. На их структуру и динамику влияют не только абиотические, но, прежде всего, биотические факторы. Плотность популяции (обилие), т.е. число (или биомасса) особей на единицу площади или объема, возрастной состав и генофонд – основные параметры экологии популяций.
Плотность популяции максимальна, когда она такова, что большая плотность уже не могла бы поддерживаться данной экосистемой, и минимальна, когда при меньшей плотности члены популяции уже не имели бы нормальных шансов на размножение (на встречу партнера).
12.4.1. Изменения плотности популяции
Если не учитывать иммиграцию и эмиграцию, плотность популяции определяется внутренними (конструктивными) факторами – рождаемостью и смертностью, а на них в свою очередь влияют внешние (регуляторные) абиотические и биотические факторы.
Общая рождаемость – это число новых особей, (Nn), добавляющихся в единицу времени (t). Так как оно зависит от числа уже имеющихся особей, лучше использовать удельную рождаемость:
![]() (12.1)
(12.1)
Идеальная (максимально возможная) рождаемость реализуется только в оптимальных условиях. Сопротивление среды (нехватка пищи, партнеров, мест для размножения, неблагоприятная погода и т.п.) сводит идеальную рождаемость к реальной (экологической).
То же, но с обратным знаком верно и для смертности. Это число особей, погибающих в единицу времени (Nm). Удельная смертность (d) получится, если отнести эту величину к численности популяции:
![]() (12.2)
(12.2)
Идеальная (минимальная) смертность проявляется как результат физиологической смертности от старости только при оптимальных условиях жизни. Сопротивление среды (погода, конкуренция, враги) повышает ее до реальной (экологической) смертности.
От баланса между рождаемостью и смертностью (b d) зависит, в какой мере изменяется плотность популяции. В идеальных условиях (при максимальной рождаемости, минимальной смертности и стабильном возрастном составе) скорость роста популяции r1=b-d всегда принимает положительные значения (b>d), так как организмы всегда дают больше способного к размножению потомства, чем было бы необходимо, чтобы заменить родителей. Поэтому популяция непрерывно растет. Специфичный для каждого вида репродуктивный потенциал (скорость размножения, биологический потенциал, или потенциальная сила размножения)
![]() (12.3)
(12.3)
при полном отсутствии сопротивления среды дает экспоненциальный рост популяции.
В природных условиях рост популяции рано или поздно прекращается из-за сопротивления среды, которое увеличивается уже из-за того, что возрастает плотность популяции. Поэтому реальная кривая роста обычно принимает сигмовидную (логистическую) форму.
После начальной логарифмической фазы она асимптотически приближается к уровню максимальной плотности популяции, т.е. к плотности насыщения (емкости среды - К), причем b становится равным d (рис. 12.10).
Размер популяции поддерживается на уровне К разными способами. У видов, живущих в эфемерных местообитаниях с высоким сопротивлением среды (большие потери от врагов), или у паразитов (малые шансы найти хозяина) репродуктивный потенциал п должен быть очень большим (r1-стратеги), чтобы они могли быстро использовать редкие или небольшие шансы. Напротив, виды, живущие в долговременных, стабильных местообитаниях, с небольшим количеством врагов или совсем не имеющие их, виды с развитой заботой о потомстве, образующие семьи или
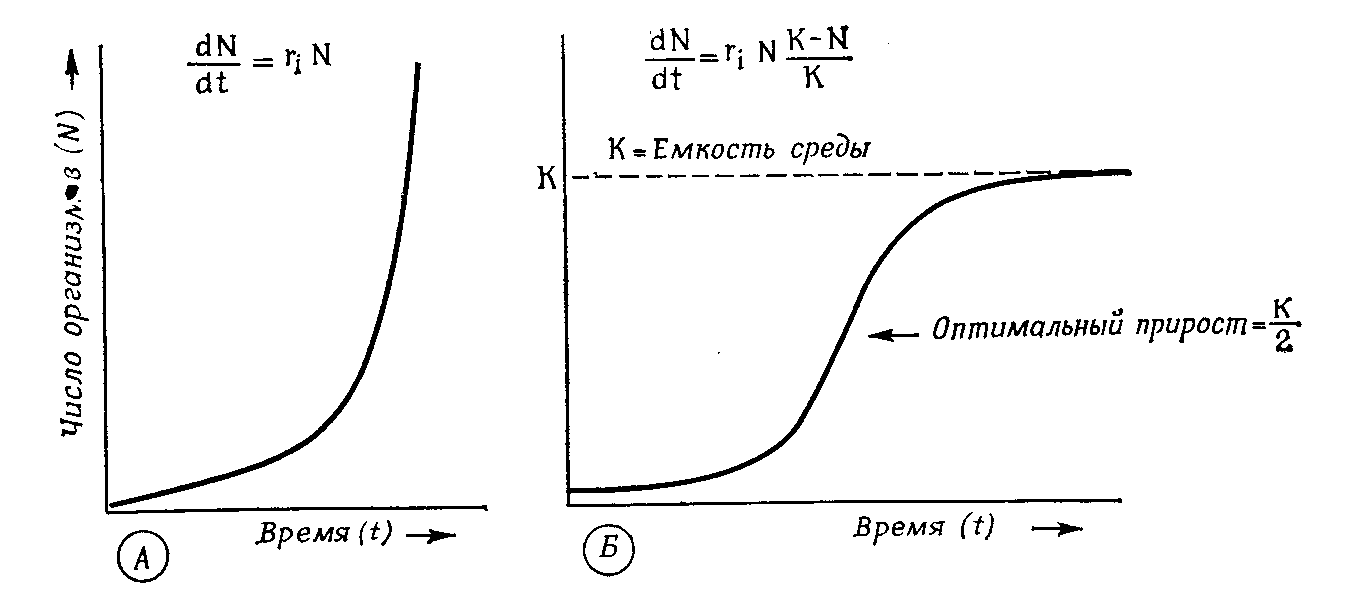
Рис. 12.10. Рост популяции. А. Экспоненциальная кривая роста при идеальных условиях среды. Б. Логистическая кривая роста в естественных условиях при емкости среды, равной К. (Wilson, Bossert)
стада, обходятся небольшим репродуктивным потенциалом (К-стратеги), так как у них значительная часть потомков достигает репродуктивного возраста. Например, орлы, дельфины и крупные копытные дают в год лишь одного потомка.