

Β-Глюкан
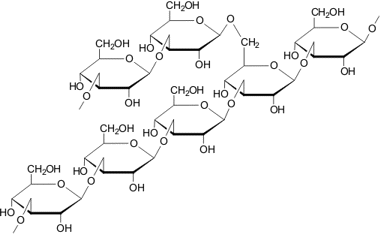
Участок молекулы β-1-3-глюкана с 1-6 боковой ветвью
Глюкан - основной структурный компонент клеточной стенкиаскомицетовых дрожжей, ответственный за поддержание ее прочности. На его долю приходится более половины общей массы клеточной стенки. Состав и строение глюкана варьирует, в его составе выделяют несколько фракций. Под термином «глюкан клеточной стенки» объединяют несколько типов полисахаридных молекул, образованных в основном β-1,3- и β-1,6-связанными остатками глюкозы. Эти компоненты различаются по растворимости в кислоте и щелочи, что дает возможность их экстрагировать раздельно.
Маннопротеины клеточной стенки
Около 40% массы клеточной стенки аскомицетовых дрожжей приходится на маннопротеины - комплекс белка с полимерами маннозы, а также небольшого количества N-ацетилглюкозамина и фосфатных групп. Маннопротеины Saccharomyces cerevisiaeразделяются на группы по характеру связи присоединения маннана к молекулам белка: между полисахаридной и белковой частью. Известны три типа таких связей: O-гликозилирование, N-гликозилирование и присоединение с помощью гликозил-фосфоинозитольного "якоря".
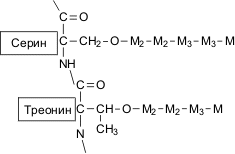
Структура О-связанных олигосахаридных цепей манопротеинов; М, М2, М3 - остатки маннозы и соответвствующие типы гликозидных связей
1. О-гликозилирование: короткие неразветвленные маннозные цепи, которые присоединены к молекуле белка при помощи О-гликозидной связи между остатком маннозы и гидроксильной группой серина или треонина. О-связанные полисахаридные цепочки содержат от одного до пяти маннозильных остатков. Количество маннозы, присоединенной к белкам таким способом может составлять около 12% от общего числа маннозных остатков в маннопротеинах клеточной стенки. Остатки маннозы связаны между собой α-1,3- и α-1,2- связями.
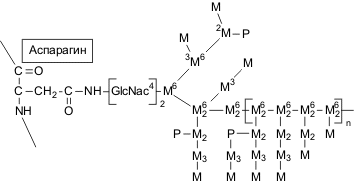
Структура N-связанных олигосахаридных цепей манопротеинов
2. N-гликозилирование: разветвленные маннозные цепи присоединяются к белку через хитобиозный мостик при помощи N-гликозидной связи между остатком N-ацетилглюкозамина и β-амидным азотом аспарагина. Высокоразветвленные маннозные цепи таких маннопротеинов неоднородны по строению и состоят из внутренней части – так называемого кора, в который входит 12–15 маннозных единиц и вариабельной по своим размерам внешней цепи. Последняя образована остатками маннозы, соединенными α-1,6-связью в центральной цепи к которой присоединены короткие олигосахариды с α-1,2- и α-1,3-связями.
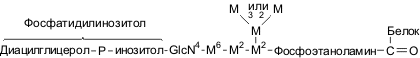
Структура GPI-«якоря» гликопротеинов клеточной стенки
3. Связь с помощью гликозил-фосфоинозитольного «якоря». Некоторые белки клеточной стенки содержат на С-конце так называемый GPI-якорь (гликозилфосфоинозитольный якорь). В отличие от N- и О-связанных цепей GPI-якорь присоединен к белку через фосфоэтаноламин нередуцирующим концом своей полисахаридной цепочки.С помощью такого «якоря» белки могут закрепляться в цитоплазматической мембране.
Хитин
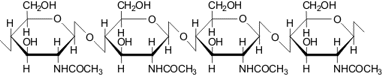
Хитин – минорный компонент клеточной стенки дрожжей. представляет собой полимер с β-1,4-связанными остатками N-ацетилглюкозамина. Примерно 90% всего хитина клеточной стенки аскомицетовых дрожжей находится в области отделения почек, в зонешрамов почкования. В этой зоне хитин участвует в построении первичной септы между материнской и дочерней клетками и образует жесткое кольцо, защищающее канал между ними. У базидиомицетовых дрожжей содержание хитина в клеточной стенке значительно выше.