

9.3 Тип Коловратки. Основные черты внешнего и внутреннего строения
Коловратки – мелкие водные животные, большей частью не превышающие своими размерами крупных инфузорий. Это преимущественно обитатели пресных вод, хотя имеются и довольно многочисленные морские представители классаRotatoria. Громадное большинство свободноподвижные, немногие ведут прикрепленный образ жизни. Известно свыше 1500 видов.
Строение и физиология. Размеры коловраток не превышают 1–2 мм, но есть среди них формы и гораздо мельче. Так, к коловраткам относится самый мелкий представитель многоклеточных Ascomorpha minina достигающий едва 0,04 мм длины.
Форма тела сильно варьирует. В редких случаях (Trochosphaera) тело шаровидно, тогда как у громадного большинства оно продолговато и разделяет участка: передний головной с мерцательным аппаратом, туловищный, содержащий все внутренности, и задний, ножной (рисунок 9.5). Нога может и отсутствовать.
Головной отдел мало обособлен от туловищного, а внутренне вовсе отделен от него. Передний конец головного отдела в некоторых случаях имеет вид диска, усаженного по краю венчиком крупных ресниц. Позади венчика на брюшной стороне головы лежит рот. Позади рта – второй венчик ресниц, более мелких. Совокупность обоих венчиков образует характерный для Rotatoria коловращательный аппарат. Реснички его находятся в постоянном мерцании, чем и обусловливают движение животного в воде, его плавание. Одновременно с этим движением ресничек в воде вызывается водоворот, привлекающий мелкие пищевые частицы ко рту коловратки.
Форма коловращательного аппарата изменчива. Так, края предротового диска могут образовать различные выросты, а за краями следует и венчик ресниц; то диск образует четыре краевые лопасти (Flosculariа), то он вытягивается в пучок длинных щупалец (Stephanoceros) и т. д.
Однако за исходную форму коловращательного аппарата всех Rotatoria в настоящее время принимают слабо дифференцированный аппарат семейства Notommatidae. У этих коловраток вместо двух венчиков мощных ресниц на брюшной стороне переднего полюса тела имеется сплошная мерцательная площадка из мелких ресничек, окружающая рот.
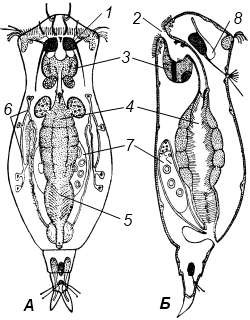
А – вид со спины, Б – вид сбоку: 1 – коловращательный аппарат; 2 – рот; 3 – мастакс; 4 – средняя кишка; 5 – задняя кишка; 6 – протонефридий; 7 – яичник; 8 – головной ганглий Рисунок 9.5 – Схема строения коловратки
При помощи особых мышц-ретракторов головной отдел может втягиваться внутрь туловищного и затем снова выворачиваться.
Туловище большею частью одето панцирем и может быть цилиндрическим, сплюснутым дорзовентрально или же сжатым с боков. Туловище содержит большую часть внутренних органов: на его заднем конце, над основанием ноги, помещается отверстие клоаки, т. е. мешка, в который впадают концевые отделы кишечника, выделительной и половой систем.
Нога – мускулистый вырост тела, одетый членистой оболочкой и снабженный на конце двумя щупальцевидными подвижными отростками– пальцами. В основании пальцев имеются две небольшие цементные железы, выделяющие особое клейкое вещество. При помощи его многие коловратки могут временно, а другие и постоянно прикрепляться к отдельным подводным предметам. В ноге хорошо развиты кольцевые и продольные мышцы. Нога позволяет коловраткам ползать, вытягиваясь по направлению движения и временно прикрепляясь передним концом к субстрату.
Стенка тела образована синцитиальным слоем гиподермы. Распространенное ранее мнение о сильном развитии кутикулы у коловраток оказалось неверным. Последние исследования, проведенные с помощью электронного микроскопа, показали, что для Rotatoria характерно наличие в гиподерме плотного волокнистого слоя под наружной цитоплазматической мембраной. Именно этому слою обязаны коловратки сохранением формы тела и из него сформированы уплотненные покровы панцирных видов. Настоящая кутикула нетипична для коловраток и известна лишь у ограниченного числа форм.
Кожно-мускульный мешок у коловраток отсутствует. В теле имеются отдельные, большей частью поперечнополосатые мышечные волокна, служащие для втягивания головного отдела, для сокращения туловища и т. д. Непосредственно под наружным эпителием лежит первичная полость тела. Пищеварительная система коловраток состоит из трех отделов. Рот лежит неподалеку от переднего конца, на брюшной стороне между венчиками ресниц коловращательного аппарата. Он ведет в ротовую полость, а та, расширяясь, – в мускулистую глотку с характерным для коловраток жевательным аппаратом, состоящим из заложенных в стенках глотки двух пар твердых зазубренных хитиновых пластинок. Это пара молоточков и пара наковален. Они трутся друг о друга, подобно паре микроскопических жерновов, и содействуют перетиранию пищи, которая состоит у коловраток главным образом из жгутиконосцев, одноклеточных водорослей и т. п. У хищных коловраток вооружение глотки способно выдвигаться через ротовое отверстие и служит для ловли добычи.
Глотка переходит в узкий пищевод, а за ним следует мешковидный энтодермальный желудок из крупных многоугольных клеток. В место соединения пищевода с желудком открывается пара желез, функция которых точно не выяснена. Желудок ведет в более узкую заднюю кишку, а последняя открывается в клоаку.
Срок пребывания пищи в кишечнике коловраток очень незначителен от времени захвата до дефекации частицы проходит от 2 до 20 мин.
Нервная система коловраток сравнительно проста, так как состоит из одного надглоточного ганглия и из отходящих от него во все стороны и кпереди, т. е. к диску, и кзади, т. е. к туловищу,нервов. Значительнее длиннее других два нерва, идущих от мозга назад по бокам кишечника через все тело до ноги. Органами чувств коловраткам служат, прежде всего, щупальца. Их чаще всего три: два брюшных и одно непарное на спине. Щупальца имеют вид маленьких конических бугорков с пучком осязательных волосков на вершине.
Большинство коловраток имеет пару или всего один глаз. Глаза лежат на самом надглоточном ганглии или же непосредственно над ним. Они устроены крайне просто: состоят из маленького лучепреломляющего хрусталика, лежащего в бокале из красного пигмента. По-видимому, это глаза инвертированного типа. Когда глаз один, то он представляет результат слияния пары глазков, обращенных друг к другу выпуклыми сторонами пигментных бокалов, почему пигмент в таком глазу дает фигуру X. Кровеносная и дыхательная системы у коловраток отсутствуют.
Выделительные органы протонефридиального типа. По сторонам от кишечника в полости тела залегают два слабо ветвящихся канальца, открывающихся сзади в клоаку. Концы веточек каналов замкнуты слепо и внутри них бьет мерцательное пламя. Число веточек варьирует от 4 до 50. Половая система. Коловратки раздельнополы и отличаются половым диморфизмом. Самки встречаются гораздо чаще самцов, и все предыдущее описание относится именно к ним. Они снабжены цельным или двухлопастным яичником, который лежит в задней части туловища, под кишечником. Обыкновенно яичник разделен на две части: производящую яйцевые клетки, или собственно яичник, и продуцирующую клетки, идущие для питания яиц, т. е. желточник. Яичник впадает при помощи короткого яйцевода в клоаку.
Самцызначительно меньше самок и часто заслуживают названия карликовых. Кишечник самцов редуцирован, выделительная система развита слабо; тотчас после оплодотворения самцы умирают. Мужская половая система состоит из единственного семенника с отходящим от него семяпроводом, впадающим в клоаку, причем конец его пронизывает мускулистый, способный к выворачиванию совокупительный орган, или cirrus. Сильная редукция многих органов у самцов объясняется кратковременностью их жизни, которая заканчивается сразу после оплодотворения самки.
Жизненный цикл и развитие.Жизненный цикл коловраток представляет собой чередование настоящих половых поколений (включают самцов и самок) с партеногенетическими, то есть имеет характер гетерогонии. Обыкновенно весной из яиц, пролежавших зиму, выходят партеногенетические самки, дающие начало опять-таки партеногенетическим самкам, и затем может следовать еще ряд таких же партеногенетических поколений (рисунок 9.6). Далее наступает половой период, когда одно из поколений партеногенетических самок начинает откладывать яйца по объему в 2– 3 раза мельче яиц, дававших до сих пор самок. Меньшим количеством материала в яйце объясняются карликовые размеры самцов и быстрота их развития. Из яиц вскоре выходят мелкие самцы, которые спариваются с самками полового, т. е. по отношению к самцам материнского поколения. Самки, которые партеногенетически производили самцов, после копуляции продуцируют особые оплодотворенные яйца. Они крупны, одеты системой плотных оболочек и называются покоящимися, ибо нормальное развитие их происходит после длительного периода покоя, длящегося от двух недель до года. Оплодотворенные яйца впоследствии дают первое поколение партеногенетических самок, с которого цикл начинается заново.
Количество жизненных циклов в течение одного года варьирует у различных коловраток: соответственно различают моноциклические, дициклические и полициклические виды. Впрочем, нередко один и тот же вид, но в разных странах или даже в разных водоемах одной и той же местности обнаруживает различное годовое число циклов. Это показывает, что цикличность коловраток в значительной мере зависит от внешних условий, меняющихся в разных водоемах и местностях. От внешних же факторов, по-видимому, зависит и появление полового поколения самок, производящих самцов. В результате значительного количества экспериментальных работ выясняется, что появление самок полового поколения обусловливается и ускоряется различными факторами: условиями питания (пониженное питание, голодание ведет к продукции самцов), химическим составом среды.
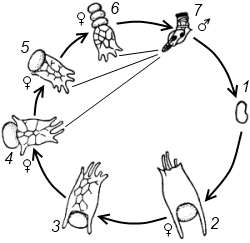
1 – яйцо (зимующая стадия); 2-6 –партеногенетические самки нескольких линий поколений; 7 – самец
Рисунок 9.6 – Годовой цикл коловратки
Цикличность коловраток усложняется наличием у многих видов сезонных изменений, не связанных с характером размножения. Так, например, у ряда следующих друг за другом в течение года поколений длинношипная зимняя форма Anuraeacochlealisможет постепенно превращаться в формы со слабо развитыми шипами и, наконец, совершенно лишается большого заднего шипа, переходя в поколение, которое называется A. cochlearistectaи раньше считалось за самостоятельный вид. К зиме она вновь возвращается к исходной форме. Такие периодические изменения в строении отдельных поколений коловраток называются цикломорфозом. Развитие яиц коловраток идет быстро; у партеногенетических яиц, не имеющих периода покоя, развитие длится 3–4 дня. Развитие прямое, без метаморфоза; в общем, дробление очень рано становится двустороннесимметричным. Другая характерная черта развития – формирование органов из небольшого числа клеток, после чего размножение клеток в них на всю жизнь прекращается. Результатом этого является постоянство клеточного состава, подобное отмеченному у представителей кл. Nematoda. Для многих коловраток точно доказано, что тело их состоит из небольшого, вполне определенного и постоянного числа клеток. Так, например, у Epiphanessenta покров состоит из 301 клетки, глотка – из 165, средняя кишка – из 76, половой аппарат – из 19, мускулатура – из 122, нервная система – из 247, выделительная система – из 24, а все тело животного – из 959 определенным образом расположенных клеток. У самцов в связи с редукцией кишечника клеток еще меньше. Постоянство клеточного состава связано с тем, что после эмбрионального периода клетки коловраток перестают делиться. С последним обстоятельством, в свою очередь, связано отсутствие у коловраток способности к регенерации. Утраченные или удаленные экспериментально участки тела не восстанавливаются. Та же особенность наблюдается и у других животных, обладающих постоянством клеточного состава, например у нематод.
Большинство коловраток живет в пресных водах, причем они делятся на донные и планктонные формы. Донные коловратки большей частью ползают или плавают у дна, но среди них имеется и целый ряд сидящих видов, последние прикрепляются выделением цементных желез ноги к субстрату, причем нога изменяется в своего рода стебелек. Большинство сидячих коловраток выделяет вокруг себя защитную трубку из прозрачного студенистого вещества или склеенных вместе в правильные комочки испражнений коловратки, которыми трубки обложены как микроскопическими кирпичами. У типичных планктонных коловраток на теле вырастают различного рода придатки, увеличивающие поверхность животного. У некоторых это длинные неподвижные шипы панциря (Notholca), у других – подвижно сочлененные с телом гребные щетинки (Polyarthra) или это особые, снабженные мышцами и усаженные щетинками выросты самого тела (Pedalion). Пресноводные коловратки играют немаловажную роль как пища различных, более крупных пресноводных организмов. Морских коловраток значительно меньше.
Небольшое количество коловраток приспособилось к жизни на суше, главным образом во влажном мху, среди лишайников и т. п. Некоторые из них (Callidina) охотно поселяются в особых мешочках на слоевище печеночных мхов, в которых долго удерживается вода.
Наземные, а также некоторые водные коловратки способны выдерживать полное высыхание, не погибая. При этом они впадают в анабиотическое состояние (мнимая смерть), но после осторожного смачивания оживают. Оживание коловраток было впервые открыто Левенгуком в 1701 г. и с тех пор многократно исследовалось. При применении ряда предосторожностей смоченные водой коловратки оживают после многолетнего высушивания. Мало того, высушенные коловратки способны в течение 4 ч выдерживать понижение температуры до –270°С (в жидком гелии) и в течение 5 мин повышение температуры до +100°С.
Коловратки играют важную роль в биологической очистке вод. Не менее важна их роль в трофических цепях в водных экосистемах.Представляют объекты питания для мальков рыб и других животных.Обычно коловраток используют при выкармливании молоди рыб в качестве добавки к основному корму при переходе с более мелких кормов к более крупным.
О происхождении коловраток существовали различные точки зрения. Долгое время их считали неотеническими личинками каких-то кольчатых червей на основании сходства Rotatoria с личинкой Polychaeta трохофорой. В последние годы установилось мнение, что сходство коловраток с грохофорой следует считать результатом конвергенции, или совпадения, связанного с одинаковым образом жизни, путь же эволюции группы Rotatoria идет от прямокишечных турбеллярий. Защитники этого мнения считают наиболее примитивными не плавающих, а ползающих коловраток и исходным типом не хорошо дифференцированные венчики, а сплошную площадку ресниц, окружающую рот. О близости к турбелляриям свидетельствует приметивное строение протонефридиев коловраток и брюшное положение рта. Из первичнополостных червей к коловраткам ближе всего гастротрихи, их сближает отсутствие кожно-мускульного мешка, наличие протонефридиев, сходство в строении мерцательного аппарата.
9.4 Тип Скребни. Строение и жизненные процессы в организме скребней Скребни – паразитические черви. Тип насчитывает около 500 видов и характеризуется следующими признаками:
Исключительно паразитические черви, в половозрелом состоянии обитающие в кишечнике позвоночных животных.
Передний конец тела преобразован в специализированный орган прикрепления в виде вворачивающегося хоботка, вооруженного рядами кутикулярных крючьев.
Покровы представлены тонкой кутикулой и гиподермой, пронизан- ной системой лакун. Полость тела первичная.
Пищеварительная, кровеносная и дыхательная системы отсутствуют. 5. Скребни раздельнополы. Выводные протоки половой системы нередко соединяются с протоками выделительной системы (устроенной по типу протонефридиев) в общий урогенитальный канал.
6. Развитие с метаморфозом. Жизненный цикл проходит со сменой животных хозяев.
В тип скребней входит только один класс с тем же названием.
Класс Скребни (Acanthocephala).Тело скребней имеет более или менее веретеновидную форму и подразделяется на собственно тело и хоботок, полый внутри и способный вворачиваться, как палец перчатки (рисунок 9.7). Хоботок усажен несколькими рядами загнутых назад кутикулярных крючьев и служит для прикрепления паразита к стенкам кишки хозяина. При помощи мышц-ретракторов хоботок вворачивается внутрь тела в особое влагалище – полый мускульный мешок, слепо замкнутый на заднем конце. Само влагалище, в свою очередь, прикрепляется к стенкам тела червя при помощи парных мышц. Тело одето тонким покровом, под которым находится гиподерма, представляющая собой синцитий, т. е результат слияния клеток эмбриональной эктодермы. Гиподерма пронизана системой каналов, или лакун. В толще гиподермы залегают немногочисленные и довольно крупные ядра. Здесь же откладываются и запасы гликогена, который является для скребней, как и для других эндопаразитов основным источником энергии.
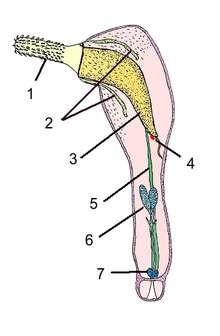
1 – хоботок; 2 – лемниски, 3 – хоботковое влагалище; 4 – ганглий; 5 – лигамент;
6 – семенники; 7 – цементная железа
Рисунок 9.7 –Схема строения самца скребня Corynosoma wegeneri
На границе между хоботком и собственно телом скребней гиподерма образует парные выпячивания, так называемые лемниски, которые в виде двух лентовидных тяжей лежат по обе стороны влагалища хоботка. Функция лемнисков до сих пор остается неразгаданной. Предполагается, что они принимают участие в процессах вворачивания и выворачивания хоботка. Кожно-мускульный мешок состоит из двух слоев: наружного – из кольцевых и внутреннего – из продольных мышечных волокон. Кнутри от мышц помещается обширная полость тела.
Пищеварительная система отсутствует в связи с паразитизмом, и питание совершается через покровы тела. По-видимому, скребни, подобно ленточным червям, воспринимают продукты пищеварения хозяина из содержимого его кишечника. Возможно, что важную роль играют при этом бесчисленные тончайшие канальцы, открывающиеся наружу микропорами, которые пронизывают поверхностный слой покровов скребней.
Нервная система очень проста. Она состоит из непарного мозгового ганглия, залегающего внутри влагалища хоботка, ближе к его заднему концу. От ганглия отходят тонкие нервные веточки вперед, к хоботку, а кроме того, два более толстых боковых нервных ствола, выходящих из влагалища хоботка, пересекающих полость тела и затем направляющихся вдоль стенок тела назад. Из органов чувств описаны лишь два небольших чувствительных сосочка – у основания хоботка и на его вершине, а также возле полового отверстия
Органы выделения найдены лишь у немногих видов. Например, у гигантского скребня (Macracanthorhynchus hirudinaceus) они имеют характер несколько видоизмененных протонефридиев. Это два древовидно разветвленных пучка, состоящих из множества тонких веточек, каждая из которых заканчивается колбовидной клеткой с мерцательным пламенем, обращенным в просвет канальца. Канальцы впадают в главные собирательные каналы протонефридиев, открывающиеся в выводные протоки поло вой системы. Таким образом, у скребней выделительная и половая системы соединены.
Половая система. Скребни раздельнополы. Половые органы парные и развитие их связано с особым соединительным тяжем, или лигаментом, который тянется от заднего конца хоботкового влагалища через полость тела к заднему концу животного. Самецимеет два овальных семенника с семяпроводами. Оба семяпровода сливаются в семяизвергательный канал, в который вливается содержимое нескольких (чаще 6) цементных одноклеточных желез. Секрет желез служит для замыкания женского полового отверстия после совершившейся копуляции. Конец семяизвергательного канала окружен мощной мускула турой и представляет совокупительный орган. Последний вдается в особый мешочек – совокупительную сумку, которая открывается наружу на заднем конце тела. Сумка может выворачиваться наружу и при копуляции охватывает задний конец тела самки.
Женская половая система у молодых особей состоит из двух яичников, залегающих внутри лигамента. С возрастом яичники распадаются на значительное количество яйцевых комков, которые сначала поступают внутрь лигамента, а затем через разрыв стенки последнего в полость тела. Каждый яйцевой комок состоит из нескольких десятков, молодых яйцевых клеток. По мере созревания яиц они оплодотворяются еще в яйцевых комках, а затем отделяются от комков и, подобно последним, плавают в полости тела. Половой проток самки устроен своеобразно. Это канал, начинающийся широко открытой в полость тела воронкой, или маточным колоколом. Помимо широкого переднего отверстия колокол на заднем конце переходит в два узких яйцевода, а на спинной стороне снабжен еше особым щелевидным отверстием, что позволяет сортировать яйца, поступающие в колокол из полости тела, а именно: узкие каналы яйцеводов пропускают только оплодотворенные, одетые оболочкой и начавшие дробиться яйца, которые имеют веретеновидную форму. Значительно более крупные незрелые яйца и яйцевые комки, попав в колокол, не проходят в яйцеводы и выбрасываются обратно в полость тела. Оба яйцевода впадают в следующий отдел полового протока – непарную длинную матку, заканчивающуюся более узким влагалищем. Влагалище открывается наружу на заднем конце животного.
Развитие и жизненный цикл. Жизненный цикл скребней обыкновенно связан с метаморфозом и со сменой хозяев. Из яйца развивается продолговатая личинка с венчиком крючков на переднем конце. Личинки и взрослые скребни состоят из сравнительно небольшого числа клеток. У некоторых скребней (Neorhynchusи др.) число клеток постоянно. Гиподерма содержит всего 6 ядер, лемниски – 3, ретракторы хоботка – 4 и т. д.; богаче всего клетками нервная система.
Для дальнейшего развития яйца с личинками должны быть проглочены промежуточным хозяином. Роль промежуточных хозяев для скребней, живущих в половозрелом состоянии в водных позвоночных (рыбах, амфибиях, в водоплавающей птице), играют главным образом ракообразные; для скребней из наземных позвоночных – это насекомые. Так, например, яйца Echinorhynchussalmonisиз сигов развиваются в рачках-бокоплавах Pontoporeia, яйца гигантского скребня из свиньи – в личинках майских жуков и бронзовок и т, д.
В кишечнике промежуточного хозяина личинка выходит из яйца, пробуравливает стенку кишки и попадает в полость тела. Здесь личинка сбрасывает крючочки, растет и постепенно превращается в почти сформированного молодого скребня; недоразвитой остается только половая система. При завершении метаморфоза происходит вворачивание хоботка во влагалище, задний конец тела впячивается, и личинка приобретшая почти шаровидную форму, окружается плотной соединительнотканной капсулой. В таком виде она может сохранять жизнеспособность неопределенно долгое время. При проматывании промежуточного хозяина позвоночным
(окончательным хозяином) его тело переваривается, и личинка скребня выходит из капсулы. Хоботок паразита выворачивается и внедряется в стенку кишки окончательного хозяина. Через некоторое время начинает функционировать половая система скребня.
У некоторых видов жизненный цикл проходит еще более сложно: так, у Corynosoma strumosum – паразита кишечника тюленя – первым промежуточным хозяином является рачок-бокоплав Pontoporeia (рисунок 9.8). Рачки эти служат пищей рыбам. В кишечнике рыб личинки скребней высвобождаются из тела рачка, проходят сквозь стенку кишки рыбы и вновь инкапсулируются в полости ее тела. Никакого развития в теле рыбы личинки не претерпевают. Они достигают половой зрелости лишь в кишечнике тюленя после поедания им зараженной рыбы. Патогенное значение скребней довольно значительно. Тюлень заражается скребнями, поедая рыбу с личинками паразита.
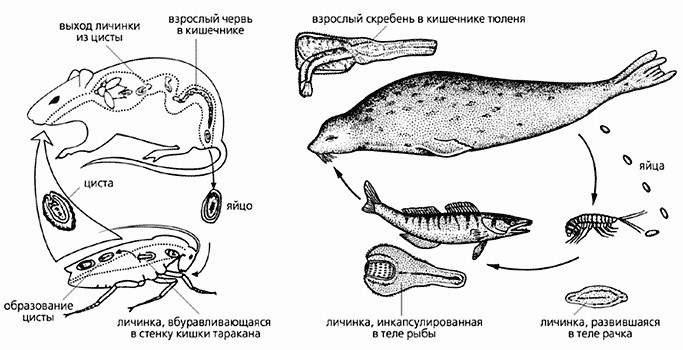
Рисунок 9.8 – Жизненные циклы двух видов скребней: один (из рода Moniliformis) паразитирует в кишечнике грызунов (слева), другой (Corynosoma strumosum) – тюленей
Гигантский скребень (Macracanthorhynchus hirudinaceus) встречается главным образом в свинье. Это крупный червь, достигающий длины 25 см и более обладающий сравнительно коротким хоботком, которым он внедряется в стенку кишки, нарушая ее целостность (рисунок 9.9). Личинки его живут в полости тела личинок бронзовок и майских жуков. Эти личинки жуков встречаются в земле, и свиньи поедают их, разрывая землю.
Некоторые виды скребней, например Pomphorhynchus laevis, вызывают серьезные кишечные заболевания пресноводных рыб. Иногда в кишечнике одного усача имеется около 300 скребней, что приводит к глубоким изъязвлениямстенки кишки. Скребни рода Polymorphus – паразиты уток – могут наносить серьезный ущерб птицеводству, так как нередко становятся причиной массовой гибели птиц, в особенности молодняка.

Рисунок 9.9 – Гигантский скребень (Macracanthorhynchus hirudinaceus)
Филогения типа Nemathelminthes. Долгое время происхождение классов типа Nemathelminthes оставалось неясным, и мнения авторов по этому вопросу сильно расходились. Лишь в последнее время, после более углубленного изучениягрупп, относящихся к этому типу, стало ясно, что все они своими филогенетическими корнями связаны с ресничными червями. В самом деле, признаки организации, общие с ресничными червями, рассеяны в разных классах типа Nemathelminthes. Так, у примитивных коловраток и брюхоресничных червей сохранились еще участки тела, покрытые ресничками. В этих классах органы выделения представлены типичными протонефридиями. Нервная система нематод (особенно у примитивных свободноживущих морских форм) и гастротрих не противоречит основной схеме строения таковой ресничных червей. Глотка первичнополостных червей, в ее более примитивных формах (классы Gastrotricha, Nematoda и Kinorhyncha), сходна с массивной глоткой прямокишечных турбеллярий. Положение рта на переднем конце тела, характерное для первичнополостных, встречается в пределах отряда Rhabdocoela. В строении половой системы, если не считать раздельнополого состояния у многих представителей типа Nemathelminthes, нет принципиальных отличий от примитивных форм из класса Turbellaria, a у коловраток женский половой аппарат снабжен желточником. Имеются, следовательно, все основания считать, что предками типа Nemathelminthes были какие-то ресничные черви.
Значительными отличиями от ресничных червей в организации первичнополостных являются наличие первичной полости тела и задней кишки с анальным отверстием. Однако эти признаки являются результатом прогрессивной эволюции и не препятствуют принятию вышеизложенной теории происхождения типа Nemathelminthes. Интересно, что у некоторых прямокишечных турбеллярий лакуны в паренхиме настолько велики, что в них могут свободно плавать паразитические инфузории. Увеличение и слияние лакун могло привести к появлению обширной первичной полости тела.
В строении отдельных классов типа Nemathelminthes имеются, на первый взгляд, значительные отличия от турбеллярий. Для первичнополостных червей характерна сильная кутикуляризация и более или менее полное исчезновение ресничного покрова, а также установление постоянства клеточного состава (коловратки и отчасти нематоды). У нематод отсутствует кольцевая мускулатура, в нервной системе образовалось окологлоточное кольцо, а протонефридии заменены выделительными каналами кожного происхождения. У коловраток выработался коловращательный аппарат; очень сильным изменениям подверглась нервная система, глотка превратилась в жевательную, произошло распадение кожномускульного мешка на отдельные мышцы и т. д. У брюхоресничных и киноринх кожно-мускульный мешок также дал начало специализированным мышцам и т. д. Все эти отличия, однако, касаются главным образом отдельных органов и тканей и мало затрагивают основной план строения.
Относительно происхождения типа Acanthocephales нет единого мнения. Чаще скребней сближают с нематодами, указывая на цилиндрическую форму тела, сходство в строении гиподермы, присутствие у них первичной полости тела и раздельнополость. Однако сближение это не имеет достаточных оснований. Тело скребней становится цилиндрическим лишь после фиксации. В отличие от нематод гиподерма пронизана лакунами, имеются протонефридии, а половая система устроена принципиально иначе. Существует и другая точка зрения, по которой скребни считаются группой, близко родственной приапулидам. В молодом состоянии тело не которых представителей класса Priapulida подразделяется на покрытую шипами голову (ее считают гомологом хоботка скребней), гладкую шейку, подобную основной части хоботка скребней, и туловище Выделительными органами у приапулид служат протонефридии, при том открывающиеся в половые протоки, подобно тому как это имеет место у скребней.
Существенные черты различия между приапулидами и скребнями объясняют паразитическим образом жизни последних.
Наиболее старая гипотеза сближает скребней с плоскими червями. В пользу этого говорит устройство хоботка, похожего на вооруженные венчиками крючьев сколексы цестод, отсутствие пищеварительной системы, органы выделения в виде протонефридиев и, наконец, усматриваемые некоторыми авторами следы ортогона в строении нервной системы скребней. Однако не следует забывать, что сходство в строении прикрепительных органов и отсутствие кишечника у скребней и цестод связано с их паразитическим образом жизни и скорее всего, носит конвергентный характер. По-видимому, следует рассматривать скребней, как самостоятельный тип животных, связанных в своем происхождении с плоскими червями, к которым они все-таки наиболее близки. Вероятнее всего, это слепая ветвь эволюции плоских червей, развивавшаяся параллельно с типом Nemathelmmthes, но независимо от него и ушедшая в сторону узкой специализации, обусловленной паразитическим образом жизни.