

7.6 Особенности симметрии, внешнего и внутреннего строения гребневиков
Тип Ctenophora, объединяющий около 100–150 видов, имеет единственный класс, носящий то же название. Научное название их типа Ctenophora происходит от греческих ktenos («гребень») и phora («носить»).
Гребневики – морские свободноплавающие, реже ползающие или сидячие радиально-симметричные животные. Размеры тела гребневиков варьируют от нескольких миллиметров до 30 см и более. Так, лентовидный венерин пояс (Cestum veneris) достигает в длину 1,5 м.
Гребневики широко распространены по всему миру и найдены во всех частях Мирового океана. Наиболее изученными остаются прибрежные и мелководные планктонные формы (роды Pleurobrachia, Beroe и Mnemiopsis) (рисунок 7.11). Гребневики, живущие у поверхности воды, обычно прозрачны и бесцветны. Глубоководные виды, напротив, имеют яркую пигментацию (например, вид «красная Тортуга» (Tortugas red).
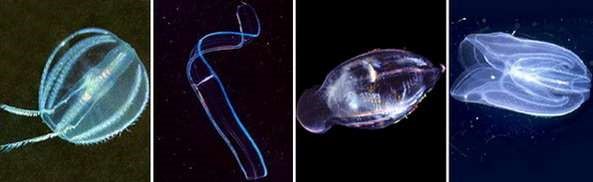
Рисунок 7.11 - Гребневики. Слева направо: плеуробрахия, венерин пояс, бероэ, мнемиопсис
Отличие от медуз:
медузы толкают себя вперед сжатием собственных тел, гребневики передвигаются за счет ритмического биения восьми своих рядов из многих тысяч волосовидных cilia (по-гречески «ресничек»),
гребневики, в отличие от медуз, не жалят,
у гребневиков длинные убирающиеся щупальца, покрытые так называемыми коллобластами – особыми клетками, которые выделяют клейкую слизь, позволяя щупальцам играть роль ловчего аппарата,
у гребневиков имеются анальные поры (у медузы непереваренная пища выбрасывается через рот),
медуза может регенерировать только утраченные щупальца, у гребневика даже половина тела способна регенерироваться в целую особь.
Симметрия тела. Симметрия тела гребневиков весьма своеобразна и является двухлучевой, то есть через тело гребневика можно провести две воображаемые взаимно перпендикулярные плоскости, каждая из которых разделит его на две идентичные половины (две плоскости симметрии). Плоскость, по которой сплющена глотка, называется глоточной, а та, в которой происходит сплющивание желудка, – щупальцевой, так как в ней же лежит пара щупалец; эти плоскости и являются плоскостями симметрии. То есть тело гребневиков построено по радиальному, а именно по двухлучевому типу. Однако следует заметить, что многие органы (ряды гребных пластинок, радиальные каналы, половые железы) расположены вокруг главной оси в числе 8. Таким образом, общее строение тела гребневиков обнаруживает сочетание двух типов симметрии: двухлучевого и восьмилучевого.
Внешняя морфология. Тело чаще всего мешковидной формы и имеет два противолежащих полюса: оральный, на котором располагается рот, и аборальный, на котором находится особый аборальный орган, похожий на нервный ганглий (рисунок 7.12). На аборальной половине тела находятся два длинных щупальца, по одному с каждой стороны. Они берут начало в глубоких карманах, выстланных ресничным эпителием – щупальцевых влагалищах. Каждое щупальце снабжено боковым рядом тонких нитевидных выростов – тентилл. Щупальца способны сильно вытягиваться и сокращаться.
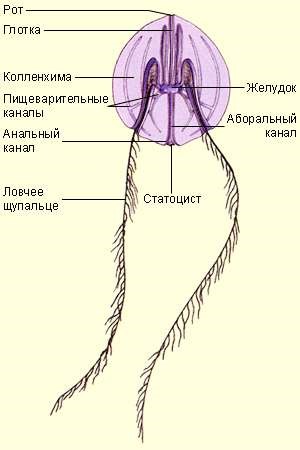
Рисунок 7.12 – Строение гребневика
Гребневики – самые крупные одиночные животные, использующие ресничный способ локомоции. От одного полюса к другому на поверхности гребневика тянется 8 продольных рядов довольно длинных ресничек (до 2 мм), которые позволяют гребневикам перемещаться со скоростью до 50 мм/с. Реснички располагаются компактными пластинчатыми группами – гребными пластинками, или ктенами, каждая из которых включает несколько тысяч ресничек. 8 рядов ктен начинаются от аборального полюса и тянутся к оральному, не доходя до ротового отверстия. Пластинки бьют по воде в одном направлении, действуя наподобие множества маленьких весел, и передвигают животное ротовым полюсом вперед. Таким образом, гребневики в течение всей жизни движутся при помощи измененных ресничек, тогда как у кишечнополостных этот способ движения имеется лишь у личинок, во взрослом же состоянии заменяется мускульным движением.В работающих гребных пластинках происходит дифракция света, из-за чего возникает эффект радуги. Тело, как и у кишечнополостных, состоит из двух слоев клеток – эктодермы и энтодермы, между которыми имеется толстый слой мезоглеи. Значительная часть объема тела гребневиков занята мезоглеей, несущей опорную функцию. Мезоглея представляет собой упругий гель, на 99 % состоящий из воды и содержащий разнообразные клетки и коллаген.
Мезоглея гребневиков хорошо развита, прозрачна и водяниста, как у медуз.
Наружный слой образован: ресничными клетками, причем их апикальный конец несет не одну, а много ресничек; интерстициальными (промежуточными) клетками, функционирующими как стволовые; секреторными (железистыми) клетками, выделяющими слизь; чувствительными клетками нескольких типов; особыми клетками, присущими только гребневикам – коллоцитами, или коллобластами.
По всей поверхности тела распределены только чувствительные и железистые клетки, другие клетки приурочены только к определенным органам (гребными пластинкам, щупальцам, щупальцевым влагалищам). В эпидермисе тентилл и щупалец имеются многочисленные специализированные клетки – коллоциты, которые выделяют клейкое вещество и приклеиваются к добыче.
Внутренний слой эпителия образован: миоэпителиальными клетками; и нервными клетками, формирующими нервную сеть. В мезоглее находятся разнообразные клетки: миоциты, нервные клетки, пигментные клетки амебоидные клетки, которые формируют основу мезоглеи.
Гребневики не имеют собственных стрекательных клеток, однако Haeckelia rubra, имеющая щупальца без тентилл и коллобластов, может вставлять в свои щупальца стрекательные клетки съеденных гидрозоев.
Мышечная система. Мышечные элементы располагаются в эпидермисе и мезоглее. В эпидермисе мышцы могут быть представлены продольными и/или кольцевыми пучками в зависимости от видовой принадлежности гребневика. При хорошем развитии обоих видов пучков формируется мышечная сеть. Мышечные элементы мезоглеи представлены продольными, поперечными или радиальными миоцитами. За счет работы мышц эпидермиса и мезоглеи у разных гребневиков могут осуществляться открывание и закрывание рта, изменение объема глотки, втягивание аборального органа и гребных пластинок.У подавляющего большинства гребневиков имеются только гладкие мышечные элементы. Поперечнополосатые мышцы обнаружены только у гребневика Euplokamis dunlapae и участвуют в раскручивании тентилл, необходимых для ловли добычи. Характерные для стрекающих эпителиально-мускульные клетки у гребневиков отсутствуют.
Внутреннее строение. Пищеварительная система гребевиков (целентерон) представлена несколькими отделами и каналами, по которым питательные вещества разносятся по телу. Рот ведет в энтодермальную глотку, имеющую вид сильно сплющенной в одном направлении трубки; кверху глотка переходит в энтодермальный желудок – мешок, сплющенный в направлении, перпендикулярном к плоскости сплющивания глотки
(рисунок 7.13).
От желудка отходят каналы гастроваскулярной системы: один направлен к полюсу, противоположному рту, и непосредственно под аборальным полюсом распадается на 4 короткие ветви: две слепые и две, заканчивающиеся узкими отверстиями. Другие два канала отходят от желудка в щупальцевой плоскости и в направлении, перпендикулярном главной оси тела, – это каналы первого порядка; каждый из них на пути к краю тела дважды дихотомически ветвится, давая 2 канала второго и потом 4 канала третьего порядка. Получившиеся таким образом 8 каналов третьего порядка впадают в соответственное число меридиональных каналов, идущих параллельно поверхности тела от аборального полюса к оральному; оба конца меридиональных каналов замкнуты слепо.
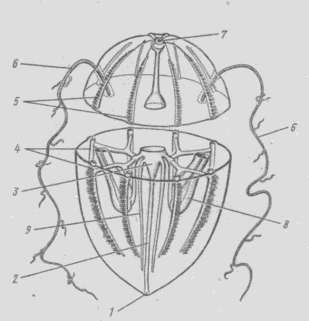
1 – ротовое отверстие, 2 – глотка, 3 – желудок, 4 – меридиональные каналы, 5 – ряды гребных пластинок, 6 – щупальца, 7 – аборальный орган, 8 – влагалища щупалец, 9 – каналы, идущие к оральному полюсу
Рисунок 7.13 – Схема строения гребневика
От желудка отходят еще два канала, идущие к оральному полюсу по сторонам глотки. Почти все известные гребневики – хищники: известен лишь один паразитоидный род Lampeia, чьи личинки паразитируют на сальпах. Представители рода Beroe питаются преимущественно другими гребневиками, большая часть нектоновых форм питается зоопланктоном разных размеров, начиная личинками моллюсков и заканчивая небольшими ракообразными наподобие копепод, амфипод и криля.
Гребневиков принято считать «тупиками» пищевых цепочек: низкое содержание питательных веществ делает их непривлекательными пищевыми объектами. Гребневиками могут питаться рыбы. Помимо рыб, гребневиками питаются медузы, морские черепахи.
Захват пищи осуществляется при помощи особых клейких клеток эпителия щупалец (рисунок 7.14). Наружная поверхность клейкой клетки имеет вид полушария, покрытого каплями липкого секрета. От основания полушария внутрь щупальца отходит спирально закрученная нить, прикрепленная другим концом к пучку продольных мышц, проходящих по оси щупальца и его ветвей.
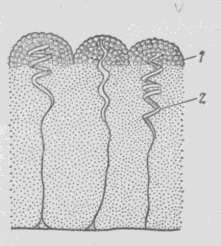
1 – полушарие с липкими сосочками, 2 – спиральная нить
Рисунок 7.14 – Строение клейкой клетки гребневик
Мелкие морские организмы при соприкосновении со щупальцами приклеиваются к клейким клеткам. Если добыча пытается вырваться, то клейкие клетки отходят от поверхности щупальца, но остаются соединенными с ним посредством эластичной спиральной нити, которая, как пружина, амортизирует рывки захваченного животного. Пойманная щупальцами добыча захватывается подвижными краями широкого рта.
Нервная система гребневиков состоит из поверхностного сплетения нервных клеток, которые под рядами гребных пластинок и мерцательными шнурами образуют более плотные тяжи, идущие к аборальному полюсу. Здесь расположен своеобразный орган чувств, получивший название аборального органа (рисунок 7.15).
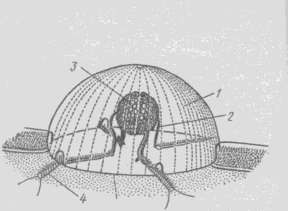
1 – колпачок из слившихся ресничек, 2 – дужки, 3 – статолит, 4 – мерцательные бороздки
Рисунок 7.15 – Схема строения аборального органа
Основу его составляет утолщение эктодермы в виде подушечки, прикрытой прозрачным колпаком, образующимся из венчика слипшихся между собой длинных ресничек. Внутри колпака над поверхностью подушечки расположены 4 крючковидные дужки, образованные длинными слившимися жгутиками, отходящими от рецепторных клеток, связанных с аборальным органом. На дужках подвешен статолит, состоящий из склеенных между собой зерен фосфорнокислого кальция.Основания дужек сообщаются при помощи мерцательных жгутов с рядами гребных пластинок. Хотя весь этот аппарат трактуется как орган равновесия, он служит также и для регуляции движения. У животного с удаленным аборальным органом движение гребных пластинок нарушается.
Органы выделения. О механизмах выделения продуктов обмена у гребневиков известно мало. Аммиак, образующийся при деградации белков, по-видимому, удаляется через поверхность тела за счет диффузии. Другие продукты обмена, скорее всего, выделяются из тела с жидкостью, заполняющей каналы целентерона, через рот или анальные поры.
Гребневики гермафродиты, причем половые клетки их дифференцируются в энтодерме. По бокам каждого меридионального канала пищеварительной системы залегает с одной стороны колбасовидный яичник, с другой – такой же семенник. Зрелые половые клетки выходят посредством прорыва разделяющей их стенки в просвет меридиональных каналов, а оттуда через рот наружу, где и происходит оплодотворение.
Жизненный цикл гребневиков простой, без метаморфоза. Дробление полное, но не равномерное, один полюс зародыша состоит из быстро дробящихся мелких микромеров, другой – из медленно делящихся макромеров. Микромеры дают эктодерму, макромеры – энтодерму. Микромеры обрастают книзу массу макромеров, которая к тому времени обнаруживает на нижнем полюсе впячивание – первичный рот и зачаток гастральной полости. Получается гаструла. По краям бластопора эктодерма впячивается и образует глотку. В глубине гастральной полости часть клеток энтодермы уходит в промежуток между эктодермой и энтодермой (т. е. в остаток полости бластулы) и дает там крестообразную группу клеток. Из этих клеток впоследствии формируются клеточные элементы мезоглеи и мускульные оси щупалец. Интересно, что здесь мы впервые видим во время развития обособленные зачатки специального третьего клеточного зародышевого пласта – мезодермы. Этим слабым зачатком является вышеупомянутая крестообразная клеточная пластинка.
Ctenophora делятся на два класса: Tentaculata и Atentaculata. Первые в течение всей жизни или только на ранних стадиях развития имеют щупальца, вторые лишены щупалец на всех стадиях. Большинство видов относится к щупальцевым (Tentaculata). Некоторые из них имеют сильно измененную форму тела, которое вытягивается в поперечном направлении (в глоточной плоскости) в ленту до 1,5 м длины, таков «Венерин пояс» – Cestus veneris.
Большой интерес представляют Platyctenidea. Эти гребневики сплющены по направлению главной оси и не только плавают, но могут и ползать по субстрату на уплощенной ротовой стороне. При переходе от плавающего к ползающему образу жизни организация гребневиков претерпевает глубокие изменения. Исключение среди них представляет арктическая форма Tjalfiella, которая ведет сидячий образ жизни. Молодь ее имеет вид типичных гребневиков. Молодое животное садится ртом на субстрат, средняя часть рта зарастает, а его концы вытягиваются кверху в трубки, служащие для захвата пищи.
К бесщупальцевым гребневикам относится обычный в северных и дальневосточных морях Beroe cucumis. Питается он другими гребневиками, заглатывая их широко раскрывающимся ртом.
Наиболее известной экологической катастрофой, связанной с гребневиками, стала инвазия вида Mnemiopsis leidyi. Вселение крупного и активного хищника – мнемиопсиса ледулы в Черное море произошло с балластными водами в танкерах от Атлантического побережья Северной Америки в 80-х годах прошлого века.
Колоссально размножился, уже в 1988 году его масса составляла около 800 миллионов тонн, то есть в среднем 1, 5 кг на 1 метр квадратный. Толерантный к малым перепадам солености и температуры воды, этот вид нанес большой урон, пожирая как зоопланктон, так и икру местных рыб. Количество планктона сразу уменьшилось в 2–3 раза. Уменьшились запасы кильки и шпрот. Самой пострадавшей стороной в этой экологической катастрофе оказалась хамса – одна из главных промысловых рыб Черного моря – ее численность резко сократилась.
Результатом, помимо прочего, стала эвтрофикация указанных водоѐмов. Положение ухудшалось отсутствием хищника, способного контролировать численность гребневика. В течении 1999-2000 годов начало отмечаться широкое распространение мнемиопсиса в водах Среднего и Южного Каспия. Найдя здесь подходящие экологические и кормовые условия, гребневик не только освоил практически всю акваторию Каспия, создав мощную популяцию с высокой численностью, но и начал влиять на всю экосистему моря. В феврале 2003 года в южной части Каспия концентрация мнемиопсиса достигала до 320 экземпляров на кубометр воды. За три года его поголовье выросло настолько, что в лунные ночи море фосфоресцировало. Именно с жизнедеятельностью гребневика ученые связывают массовую гибель каспийской кильки летом 2001 года. По данным специалистов Дагестанского отделения Каспийского научно-
исследовательского института рыбного хозяйства, тогда погибло порядка 200 тыс. тонн кильки, что составляло пятую часть ее общего количества в бассейне Каспия. Далее по пищевой цепочке наступила массовая гибель каспийского тюленя. При этом в первую очередь популяция теряла весь ожидаемый приплод (не нагулявшие жир животные либо не вступали в размножение, либо рожали ослабленных детенышей, которые вскоре гибли).
В данное время популяция кильки снизилась на порядок, а следом произошло и снижение количества осетровых рыб. Вдобавок мнемиопсис пожирает их икру, не давая шансов на размножение. Уменьшению популяции M. leidyi способствовала интродукция североамериканского хищного гребневика Beroe ovata и локальное похолодание 1991–1993 годов, однако восстановлению до первоначального уровня популяция зоопланктонных организмов в указанных морях, скорее всего, не подлежит.
В 2006 году Mnemiopsis появился на востоке Средиземного моря и сейчас угрожает Балтийскому и Северному морям. Некоторые виды образуют массовые скопления в пелагиали и забивают рыбацкие сети, мешая прибрежному рыболовству (например, Pleurobranchia pileus в Северном море).
Гребневики могут выступать в роли стабилизаторов морских экосистем, регулируя численность зоопланктона.
Филогения типа Ctenophora. Гребневики обладают некоторыми примитивными чертами, например движением при помощи измененных ресничек. Развитие гребневиков обнаруживает следы закладки мезодермы – этим они приближаются к червям. Возможно, что гребневики отделились от общего с Coelenterata ствола развития до того, как последние стали сидячими животными. Следует отметить, что наличие у гребневиков двух различных плоскостей симметрии, одной пары щупалец и некоторые другие признаки указывают на известное изменение данной группы в сторону перехода к билатеральной симметрии.