

31.3 Современные палеонтологические данные и место архиоптерикса в эволюционном древе птиц
Птицы являются «дальними родственниками» пресмыкающихся. Скорее всего, отделение птиц от группы пресмыкающихся, бывших предками крокодилов, динозавров и летающих ящеров, случилось в конце триасового или начале юрского периода мезозойской эры (т. е. около 170–190 млн. лет назад).Первые птицы появились в юрский период (между 201 и 145 млн. лет назад).
Проблема заключается в отсутствии предков птиц или «первой птицы».
Найденные отпечатки трактуют по-разному и ни одну находку однозначно не считают предком современных птиц. Прямые предки птиц не обнаружены до сих пор.Согласно традиционным представлениям эволюция птиц началась в юрском периоде. Согласно этой версии, самым древним из известных видов птиц считается археоптерикс (Archaeopteryx lithographica) (рисунок 31.1).
Птицы в этом случае рассматриваются как специализированные представители теропод, близкие к другим оперенным динозаврам из группы манирапторов, например дромеозавридам.
Археоптерикс. Самая первая находка, обнаруженная в Баварии в 1861 году. По обнаруженным отпечаткам было описано небольшое существо величиной с ворону, жившее около 150 млн. лет назад. На принадлежность к птицам указывает наличие перьев. Анатомически больше похож на рептилию. Полноценно летать не умел. Возможно, только планировал с ветки на ветку. Однако археоптерикс отнесен к классу Птицы, подклассу Ящерохвостые.

Рисунок 31.1 – Археоптерикс – самая древняя птица
Энанциорнис. Останки древних птиц были обнаружены в Аргентине в 1981 году. Жили 70-65 млн. лет назад и обладали признаками птиц: имели хорошо развитые крылья, умели летать. Наличие зубов и строение скелета роднят находку с археоптериксом.
Конфуциусорнисы. Самая древняя птица, независимо утратившая зубы, была найдена в Китае. Обитала около 120 млн. лет назад. Клюв был покрыт роговым чехлом. По отдельным признакам скелет схож с современными птицами.
Дромеозавриды (рисунок 31.2). Обитали в период между 168 и 66 млн. лет назад. Это обширное семейство, относящееся к подотряду Тероподы, отряду Ящеротазовые, содержит несколько видов динозавров с перьями (дейноних, ютараптор, синорнитозавр). Наиболее значимым является микрораптор или «четырехкрылый динозавр», имевший крылоподобные поверхности на передних и задних конечностях.

Рисунок 31.2 – Дромеозавриды
Анхиорнис (рисунок 31.3). Найден и описан в Китае в 2009 году. Относится к семейству Троодонтиды, отряд Ящеротазовые. Обитал 167-155 млн. лет назад. В длину достигал 30-40 см и весил 100 г. Имел оперение, длинный хвост, клюв.
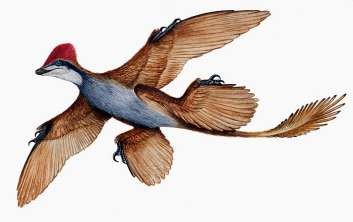
Рисунок 31.3 – Анхиорнис
Существуют и другие формы, указывающие на наличие перьев у динозавров. Например, каудиптерикс, живший 120-125 млн. лет назад, имел на хвосте веерные перья, служившие, вероятнее всего, для привлечения полового партнера.
Найденные формы указывают на попытки разных видов освоить воздушное пространство. От какой именно линии произошли птицы, еще предстоит узнать.
Анализ находок позволил сформировать гипотезу происхождения и эволюции птиц от предка-динозавра. Долгое время считалось, что троодонтиды и дромеозавриды – «пернатые динозавры» – являются ближайшими предками современных птиц.
Эволюция именно этих пресмыкающихся пошла путем осваивания лазания по деревьям, из-за чего задние конечности использовались для опоры тела, а передние понемногу переквалифицировались для обхвата ветвей (пальцами). В дальнейшем появилась способность этих животных к порханию с ветки на ветку, а затем – к планирующему полету.
На основе исследований экземпляров выдвигались различные гипотезы филогенетических отношений птиц, но общепринятой филогении нет.
Классические гипотезы: «с деревьев вниз»
Текодонтная гипотеза. Эту гипотезу предложил в начале XX века Герхард Хайльманн, основываясь на детальном обзоре биологии птиц (включая археоптерикса) и рептилий. По его мнению, наиболее близкими родственниками птиц следует считать триасовых псевдозухий (из надотряда текодонтов). Текодонтная гипотеза была общепринятой в середине XX столетия и поддерживается сегодня рядом специалистов.
Однако вплоть до настоящего времени среди псевдозухий не удалось обнаружить форм филогенетически близких к археоптериксу или другим ранним птицам.
Гипотеза Уокер (архозавроморфная). Уокер обосновывал в 70-ых годах XX века родство археоптерикса и всех птиц с триасовыми архозаврами из подотряда сфенозухий, родственных современным крокодилам. Аргументы Уокера неоднократно критиковались, и сам автор впоследствии признал гипотезу несостоятельной. До недавнего времени гипотезы постепенно теряли сторонников. Однако, после открытия четырехкрылого оперенного динозавра в Китае, предсказанного еще Бибом в 1915 году под именем «тетраптерикс» (Tertrapteryx), группа гипотез «с деревьев вниз» вновь переживает расцвет.
Гипотезы: «с земли вверх»
Тероподная гипотеза. Гипотеза первоначально была выдвинута Томасом Генри Хаксли в XIX веке. В 70-ых годах XX века ее возродил Джон Остром. Основываясь на сравнении пересмотренной им остеологии археоптерикса и тероподных динозавров (в особенности описанного им дейнониха), Остром заключил, что ближайшими родственниками археоптерикса являются целурозавры из семейства дромеозаврид. В последствии выводы Острома были подтверждены: кладистическими исследованиями,обнаружением экземпляров целурозавров с отпечатками перьев, сравнением микроструктуры яичной скорлупы и репродуктивной биологии целурозавров и птиц. Сегодня тероподная гипотеза принимается большинством палеонтологов.
Однако после открытия четырехкрылого оперенного динозавра в Китае основная идея гипотезы – «с земли вверх» – стала все чаще подвергаться острой критике. При этом надо заметить, что остальные находки оперенных динозавров в Китае довольно хорошо вписываются в концепцию «с земли вверх». Вместе с тем ряд авторов придерживается альтернативной версии, согласно которой, первой достоверной находкой птиц является довольно спорное ископаемое протоавис из позднего триаса (открытие протоависа принадлежит Санкару Чаттерджу в 1984 году). Согласно этой версии, и птицы, и тероподы произошли от одной из более ранних групп архозавров.В настоящее время выдвинуто несколько различных гипотез происхождения и эволюции птиц, но общепринятой филогении нет.
У слаженной «динозавровой» гипотезы появились противники, когда в 1991 году в Техасе Шанкар Чаттерджи нашел двух ископаемых птиц – протоависов, живших 220-200 млн. лет назад, т.е. на 50-70 лет раньше археоптерикса. В отличие от «баварской птицы» протоавис имеет больше общих черт с современными птицами. Это значит, что тероподы, жившие позже протоависа, являются в лучшем случае «братьями», а не прямыми предками птиц.
Гипотеза Чаттерджи. Считал, что древнейшей птицей является протоавис, а не археоптерикс. При этом ближайшими родственниками птиц являются триасовые цератозавры – группа тероподных динозавров, обладавшая рядом существенных особенностей, характерных для птиц (близкое к гетероцильному строение шейных позвонков, сплав тазовых костей и другие). Эти особенности скелета цератозавров традиционно расцениваются как результат конвергенции. При этом полет появился в результате планирования дромерозаврид с деревьев.
Гипотеза дифилитического происхождения птиц. Данную гипотезу,которая частично основывается на изучении протоависа, активно поддерживал палеонтолог Евгений Курочкин из Палеонтологического института РАН. Согласно этой гипотезе, веерохвостые птицы произошли от некоторой группы юрских архозавров, близким родственником которых является протоавис. Ящерохвостые птицы (например, археоптерикс и энанциорнисы) являются потомками тероподов.При этом предки птицы обитали не на настоящих деревьях, а в кустарниках, на которые они запрыгивали, используя силу ног, а спрыгивали, замедляя прыжок быстрыми махами оперенных передних конечностей. Ключевым в этой гипотезе является отсутствие стадии планирующего полета, которая неоднократно критиковалась сторонниками гипотезы «с земли верх», согласно мнению которых, планирующий полет самодостаточен и не способен перейти в машущий.
Тем не менее, согласно Курочкину, планирующий полет все же имел место, но относился не к настоящим птицам, а к тупиковой ветви развития, представленной археоптериксом. Другим ключевым моментом в гипотезе Курочкина является то, что, в отличие от авторов классических гипотез «с деревьев вниз», он считает, что передние конечности при лазаньи играли вспомогательную роль и не обхватывали ствол дерева, а цеплялись за мелкие соседние ветви, помогая балансировать. Основная роль при лазаньи в гипотезе Курочкина отводится сильным задним конечностям. Это является вторым ответом критикам гипотезы «с деревьев вниз», по мнению которых, лазанье по деревьям с обхватом передними конечностями ствола дерева несовместимо с машущим полетом.
Гипотеза Диала – выдвинута в 2003 году на основе наблюдения птенцов кекликов, взбиравшихся по наклонным поверхностям (вплоть до почти вертикальных), помогая себе взмахами крыльев.
Гипотеза Гарнера «атакующий проавис» («prouncing proavis») – выдвинута в 1999 году, отвергает как идею «с земли вверх», так и «с деревьев вниз». Согласно этой гипотезе, полет появился в результате прыжков на добычу из засады с возвышения, а появление перьев служило повышению точности прыжка. В подтверждение гипотезы Гарнер приводит кладистическую линию Sinosauropteryx-Protarchaeopteryx-CaudipteryxArchaeopteryx-Iberomesornis-Enantiornithes-Ornithurae, годом ранее построенную Джи.
Гипотеза Петерса – выдвинута в 2002 году как развитие идеи Гарнера. В качестве одного из доказательств своей гипотезы Петерс раскритиковал как классическую гипотезу «с деревьев вниз», так и классическую «с земли вверх». Так, например, по его мнению, приспособление передних конечностей к лазанью противоречит приспособлению к полету.
Гипотеза Лакасы-Руиса (Lacasa-Ruiz, 1993) и Лопарева (1996) – изначально выдвинута Лакасой-Руисом в 1993 году, а затем доработана в 1996 году С. А. Лопаревым, который в качестве доказательства привел сравнительный анализ строения таза птиц, а также онтогенез современных птиц. Согласно гипотезе, предки птиц обитали на заросшем мелководье, и перья на передних конечностях изначально служили для создания тени с целью привлечения рыбы (так охотятся некоторые болотные птицы). При спасении от хищников махи передними конечностями облегчали бег по заросшему болотистому мелководью, что постепенно перешло в использование экранного эффекта, затем в транзитный полет (cruising flight).
Гипотеза Савельева – выдвинута в 2006. Согласно этой гипотезе, птицы изначально обитали на мелководье, а затем научились нырять, используя передние конечности подобно оляпке и пингвину, что явилось преадаптацией к полету.
Гипотеза Острома (Ostrom) – выдвинута в 1976 году, и согласно ей, перья на передних конечностях изначально использовались в качестве ловчих сетей для насекомых. Гипотеза неоднократно критиковалась, и в 1997 году Остром от нее отказался.
Судя по современным палеонтологическим данным, в течение мезозоя полет (как активный, так и планирующий) осваивают несколько разных групп древних рептилий. При этом классические, давно известные мезозойские летающие пресмыкающиеся – птерозавры не являются предками птиц. Широко известное птицеподобное существо из средней юры Европы – археоптерикс – относят к особому подклассу ящерохвостых птиц и не рассматривают в качестве непосредственного предка для остальных – веерохвостых – птиц. Отсутствие киля на грудине, пневматизации костей и пряжки в скелете кисти делали его вряд ли способным к активному машущему полету. Ряд признаков археоптерикса являются уникальными: например – попарное отхождение рулевых перьев от длинного хвостового отдела позвоночника.
Другие признаки птиц унаследованы от рептилийных предков – челюсти, несущие зубы, длинные пальцы кистей, несущие когти, и др. По сути дела единственным собственно птичьим признаком археоптерикса были перья, отпечатки которых хорошо видны на окаменевших остатках.
Однако в настоящее время считается общепринятым, что развитие оперения и ряда других признаков у предков птиц проходило первоначально вне всякой связи с полетом, а в качестве элементов формирующейся системы гомойотермии.
В последние десять лет проблема происхождения птиц вновь стала предметом горячих дискуссий и непримиримых противоречий, что связано как с появлением новых и обширных материалов по ископаемым птицам, так и с триумфальным шествием кладистики (или филогенетической систематики) – новой методологии изучения отношений между организмами. За последнее время число известных видов древнейших мезозойских птиц, существовавших между 145 и 65 миллионами лет назад, возросло за 20 лет с 40 до 100.
В настоящее время конкурируют 2 основные гипотезы: птицы произошли от тероподных диназавров группы манираптор или же от целурозавров в юре; птицы произошли от мелких текодонтов псевдозухий в триасе. Возможны и другие версии обоих гипотез.
Рисунок 31.4 – Struthiomimus – представитель семейства страусоподобных динозавров (Ornitomimidae) из верхнего мела
В последние десятилетия двадцатого века «под подозрение» палеонтологов попала группа некрупных страусоподобных динозавров – орнитомимов (авимимов) (рисунок 31.4), для которых характерно развитие пряжки, что можно объяснить только необходимостью единомоментного разворачивания перьев, связанных с кистью.
Предполагается, что «аплодирующие» движения оперенных передних конечностей, при помощи которых они, возможно, выпугивали насекомых из травы, способствовали развитию больших грудных мышц и грудины. При этом формирующееся крыло использовалось первоначально не для полета, а для маневров при преследовании добычи, подскоках, движении по скалам и т.п.