

Янко Слава (Библиотека Fort/Da) || http://yanko.lib.ru |
182 |
полимеры находятся в водной среде, где весьма вероятен их гидролиз.
Полимеры не имеют кристаллической структуры и состоят из большого числа повторяющихся звеньев — органических макромолекул. Каталитические реакции и процессы играют особую роль в образовании макромолекул. Термин «полимер» введен в
1833 г. шведским химиком Й. Берцелиусом (1779 — 1848). Наряду с полимеризацией могут идти и процессы деструкции — разрушение полимеров, распад их до мономеров. Оба процесса происходят как самопроизвольно, так и направленно в зависимости от условий протекания реакции и возможного воздействия на нее.
12.2.2. Аминокислоты
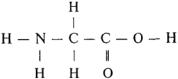
Важнейшим для биохимии соединением является аминокислота. Аминокислоты — это такие органические соединения, которые служат основным элементом построения растительных и животных белков и поэтому играют особую роль в жизни организмов, участвуя в обмене азотсодержащих веществ. Свободные аминокислоты, например, моноаминокарбоновые (глицин), содержат карбоксильную группу (—COOH) и аминогруппу (—NH2), присоединенные к одному атому углерода С:
Они обладают свойствами и кислот, и оснований. Напомним, что кислоты — химические соединения, характеризующиеся диссоциацией в водном растворе с образованием ионов водорода Н+, а основания — диссоциацией с образованием иона ОН-. Замена атома водорода в кислоте металлами образует соли, а растворимые в воде основания являются щелочами.
Природных аминокислот свыше 170, однако только 20 из них служат мономерными звеньями цепей, из которых построены полимерные молекулы всех белков. Образующиеся при удалении атома водорода от аминокислоты, аминокислотные остатки соединяются с ней пептидной связью ( —СО—NH— ), которая соединяет аминогруппу одной кислоты (или остатка) с карбоксильной группой другой. Полипептидная цепь образуется в ре-
306
зультате повторения актов пептидной связи. На одном конце этой цепочки (N-конце) свободная NH2 группа, на другом (С-конце) — COOH карбоксильная группа.
Поэтому аминокислотные остатки являются полимерами полипептидов. Если заменить хотя бы один атом или одну аминокислоту в полипептиде, то возникает новая молекула с другими свойствами и структурой. Каждый остаток имеет постоянную часть и вариабельные концевые или боковые части, через которые осуществляется пептидная связь таким образом, что формируется непрерывная основная цепь белковой молекулы. Каждая аминокислота имеет свое значение — отрицательный логарифм концентрации Н+, характеризующий кислотность рН, при котором аминокислота электрически нейтральна. При увеличении рН водного раствора аминокислоты выступают как доноры положительных ионов водорода Н+, а при уменьшении — как их акцепторы. Химические связи в цепях полипептида могут иметь разный характер. Между положительно и отрицательно заряженными боковыми группами аминокислот или их остатками возникает ионная связь, могут образоваться также водородные и ковалентные связи, что и определяет многофункциональность и значение аминокислот при построении белков живого организма. Аминокислоты входят также в состав других макромолекул — нуклеиновых кислот.
12.2.3. Теория химической эволюции в биогенезе
Проблема самоорганизации сложных макромолекулярных структур имеет общие корни с проблемой самосборки белков живых организмов: за очень короткое время полимерная цепочка из определенных аминокислотных звеньев точно складывается в определенную молекулярную конструкцию, формируя именно данный конкретный белок. Таким образом, процессы химической самоорганизации органических молекул играют важнейшую роль в предбиологической эволюции живых организмов. В настоящее время в химических науках сформировалось новое направление — эволюционная химия — наука о самоорганизации и саморазвитии химических систем, в
Горбачев В. В. Концепции современного естествознания:—М.: ООО «Издательский дом «ОНИКС 21 век»: ООО «Издательство «Мир и Образование», 2003. — 592 с: ил.
Янко Слава (Библиотека Fort/Da) || http://yanko.lib.ru |
183 |
основе которой лежат представления о решающей роли катализа при переходе от химических систем к биологическим.
В 1964 г. А.П. Руденко была предложена теория химической эволюции и биогенеза, где было показано, что эволюционирую-
307
щими элементами в развитии предбиологических химических систем являются те структуры и органические соединения, которые усиливали активность и селективность действия катализаторов. Таким образом, и на этой стадии эволюции природы происходил отбор наиболее «нужных» ей веществ для создания живых организмов. Самопроизвольная автокаталитическая реакция в природе «служит» делу эволюции, является как бы «орудием» отбора наиболее прогрессивных эволюционных изменений катализаторов. В этом смысле биокатализ с участием ферментов связан с проблемами биогенеза и происхождения жизни.
Вбиологии роль таких катализаторов выполняют специфические белковые протеины
—ферменты. Эти макромолекулы ферментов обладают такой пространственной конфигурацией, которая дает возможность изменить скорость реакции. Молекулы веществ, участвующих в реакциях, стремятся присоединиться к активным участкам молекул фермента, повышая вероятность их столкновения и, следовательно, начала химической реакции. Самоуправлением в химических процессах как раз и занимается автокаталитическая реакция: продукт этой реакции начинает управлять ее скоростью и даже самой возможностью протекания.
12.2.4. Теория молекулярной самоорганизации М. Эйгена
Другой моделью добиологической фазы образования макромолекул и их эволюции является теория молекулярной самоорганизации М. Эйгена (р. 1927), которая позволяет применить физические представления теории информации, кибернетики и термодинамики к эволюции живых организмов. В этой модели организм рассматривают как открытую термодинамическую систему, которая обменивается веществом, энергией и информацией с окружающей средой. Эти условия наряду с обеспечением за счет такого обмена стационарного неравновесного состояния системы являются причиной процесса ее самоорганизации. Живая открытая система пытается получить от окружающей среды именно свободную энергию и отдать ей «омертвленную» для него связанную энергию — энтропию, которую нельзя превратить в работу для процессов, происходящих в живом организме.
Обмен веществом (в биологии это называется метаболизмом) и информация связаны между собой. Метаболизм можно также рассматривать как совокупность всех химических реакций в живой клетке, приводящих к необходимому для нее обмену веществ. Молекулы-мономеры, переходя из внешней среды в орга-
308
низм, несут информацию, которая внутри организма перерабатывается и закрепляется при процессах полимеризации и деструкции, т.е. первоначальная информация об окружающем мире для живого организма закодирована химическим путем. Например, «память» макромолекул — это фиксированная последовательность химических связей. Полимеризация идет путем самоинструктируемой репродукции (воспроизведения) образованных макромолекул. Если скорость репродукции больше скорости распада биополимеров, то макромолекулы растут, в противном случае — распадаются.
При этом необходимо учитывать потоки поступающих мономеров от внешней среды в организм и самовоспроизведение в нем макромолекул. Такие процессы описывают нелинейными кинетическими уравнениями переноса и взаимодействия молекул. Внешние ограничения и потоки создают в системе отбор, что позволяет рассматривать селекционную ценность информации для физики эволюции живого. По Волькенштейну, «вымирают» те макромолекулы, которые не обладают максимальной селекционной ценностью.
Физическое понимание количественного представления принципа естественного отбора Дарвина и будет определяться ценностью информации, которую И.И. Шмальгаузен [153] сравнивал со степенью ее незаменимости для живого организма. Заметим, что естественный отбор имеет место и в неживой природе. Более сложные органические молекулы являются более стойкими к излучению, чем простые, поэтому в природе происходило накопление молекул более сложных конфигураций.
Такой подход вносит физический смысл в дарвиновское определение «наиболее
Горбачев В. В. Концепции современного естествознания:—М.: ООО «Издательский дом «ОНИКС 21 век»: ООО «Издательство «Мир и Образование», 2003. — 592 с: ил.