

3.6.5 Способность пирамидных нейронов поля са1 генерировать пачечные разряды сопровождается повышением возбудимости этих клеток
Полагая, что способность пирамидных нейронов поля СА1 отвечать множественным спайком на одиночный стимул после эпизодов повышения внеклеточной концентрации калия, является формой пластичности (возможно патологической), мы решили исследовать механизмы этого феномена.
Мы разделили передачу сигнала в поле СА1 на три этапа: передача возбуждения по пресинаптическим волокнам, синаптическая передача и генерация потенциала действия в пирамидных нейронах.
Для оценки возбудимости пресинаптических волокон мы построили зависимости амплитуды потенциала волокон (мВ) от силы тестирующего стимула (μА) (Рис 3.6.4а). Периодические повышения внеклеточной концентрации калия не оказывали значительного влияния на характер этих кривых. Этот результат указывает на то, что коллатерали Шаффера не меняют своей возбудимости при данном воздействии.
Для оценки эффективности синаптической передачи мы построили зависимости наклона пВПСП (мВ/мс) от амплитуды потенциала волокон (мВ). Эпизоды повышения калия так же не оказывали существенного влияния на глутаматергическую синаптическую передачу (Рис. 3.6.4б).
Для оценки возбудимости пирамидных нейронов мы построили зависимость амплитуды популяционного спайка (мВ) от наклона пВПСП (мВ/мс). Обнаружилось, что использование трех эпизодов повышения внеклеточной концентрации калия приводит к долговременному (сходного со временем генерации множественных спайков) сдвигу этой зависимости в область более высоких значений, то есть, потенцирует передачу сигнала в этом участке (Рис. 3.6.4в). Амплитуда популяционного спайка после воздействия была 248 ± 37 % от его амплитуды до калиевых эпизодов (данные получены для наклона пВПСП = 0,5 мВ/мс через час после воздействия; р=0,038).

Рис. 3.6.4 Развитие пачечной активности сопровождается увеличением возбудимости пирамидных клеток
а, Зависимость амплитуды потенциала пресинаптических волокон (коллатералей Шаффера; пВ) от амплитуды электрического стимула. Данные усреднены по нескольким экспериментам (n=5). Периодические эпизоды повышения внеклеточной концентрации калия (3 эпизода по 30 секунд с интервалом 10 минут, концентрация 20 мМ) не приводили к изменениям в возбудимости терминалей (-○- и -□- - зависимости до и через час после воздействия, соответственно).
б, Зависимость наклона (мВ/мс) полевого ВПСП (глутаматергическая синаптическая передача; пВПСП) от амплитуды потенциала пресинаптических волокон. Данные усреднены по нескольким экспериментам (n=5). Периодические эпизоды повышения внеклеточной концентрации калия (3 эпизода по 30 секунд с интервалом 10 минут, концентрация 20 мМ) не приводили к изменениям в эффективности глутаматергической синаптической передачи (-○- и -□- - зависимости до и через час после воздействия, соответственно).
в, Зависимость амплитуды популяционного спайка, регистрируемого вstr.pyramidale(возбудимость пирамидных клеток; ПС) от наклона полевого ВПСП. Данные усреднены по нескольким экспериментам (n=5). Периодические эпизоды повышения внеклеточной концентрации калия (3 эпизода по 30 секунд с интервалом 10 минут, концентрация 20 мМ) приводили к значительному повышению возбудимости нейронов (-○- и -□- - зависимости до и через час после воздействия, соответственно).
Разбросы указывают ± с.о.с

Возникает вопрос, играет ли роль это увеличение возбудимости пирамидных нейронов в их способности генерировать пачечный разряд? Возбудимость мембраны определяется способностью генерировать потенциал действия на деполяризацию. Другими словами, повышение возбудимости внесинаптической мембраны это повышение вероятности генерации потенциала действия. Способность генерировать пачечный разряд может определяется длительностью деполяризации при неизменной вероятности генерации одиночного потенциала действия. Несомненно, повышение возбудимости нейронов должно играть роль в добавлении дополнительных разрядов на затухающей фазе деполяризации (Рис. 3.6.5). Поэтому следующей задачей представленной работы был поиск возможных фармакологических способов доказательства сходства или различия механизмов генерации вызванных множественных спайков и потенциации амплитуды популяционного спайка.
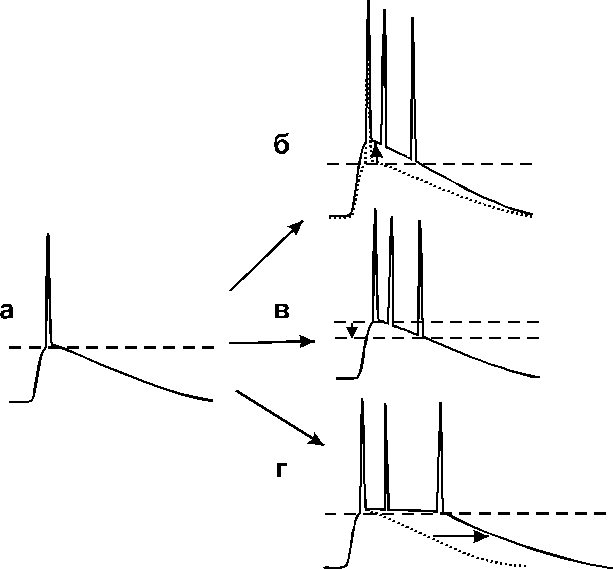
Рис. 3.6.5 Механизмы формирования пачечной активности
а, Деполяризация, вызванная ВПСП, приводит к генерации потенциала действия в нейронах (порог генерации обозначен пунктирной линией)б, ВПСТ увеличивается благодаря синаптической потенциации или облегчению электротонического проведения из дендритов в аксонный холмик. Возникает множественный разряд.в, Порог генерации потенциалов действия снижается, что приводит к генерации множественного разряда на ту же амплитуду ВПСТ.г, Множественный разряд возникает при неизменном пороге и амплитуде деполяризации, более длительной по времени.