

3.4.4 Спилловер глутамата активирует аксональные каинатные рецепторы
Полученные в предыдущих разделах результаты демонстрируют эффект аппликации экзогенного агониста каинатных рецепторов. Однако, из них не ясно может ли эндогенный глутамат, высвобождаемый синаптически, приводить к активации аксональных каинатных рецепторов. Для ответа на этот вопрос мы воспользовались протоколом аналогичным тому, который использовался для исследования гетеросинаптической депрессии ТПСТ, опосредованной метаботропными рецепторами. Мы стимулировали коллатерали Шаффера серией стимулов (5 стимулов при частоте 5 Гц), которая должна была вызвать значительный
контроль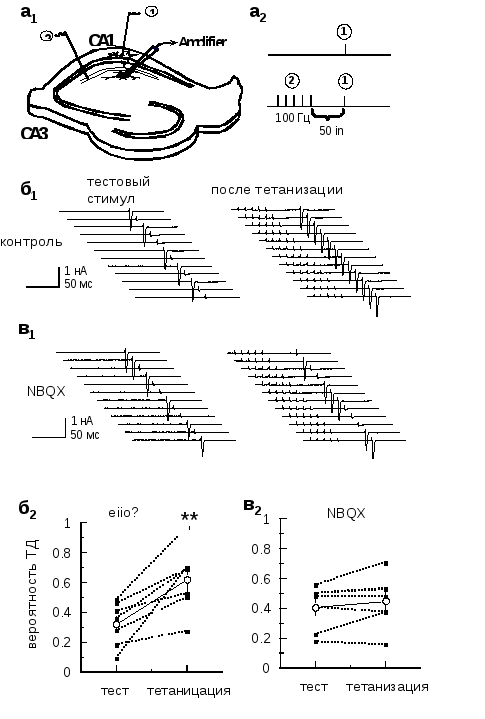
Рис. 3.4.4 Синаптически высвобождаемый глутамат увеличивает возбудимость аксонов через каинатные рецепторы
а, Экспериментальный протокол. Позиции стимулирующих и регистрирующего электродов показаны на рисункеа1. Тестовый стимул, инициирующий антидромный ТД (а2) подавался через электрод (1) либо сам по себе, либо после серии стимулов (5 стимулов при частоте 100 Гц) через дистальный электрод (2).б, Оригинальные записи, полученные в одном интернейроне, показывают, что вероятность генерации ТД увеличивается после высокочастотной стимуляции глутаматергических терминалей. Это увеличение подавлялось при блокаде каинатных рецепторов с помощью NBQX (100 μМ).в, Данные, усреденные по 7 интернейронам. **: p<0,01
выброс глутамата. Затем с интервалом 50 мс ты тестировали вероятность возникновения антидромного ТД в ответ на стимуляцию аксона в str.orience(Рис 3.4.4а). Поскольку AMPA, NMDA, ГАМКА, ГАМКBи mGluR группы III рецепторы были блокированы, мы предположили, что возможные эффекты высокочастотной стимуляции возбуждающих терминалей будут опосредованы каинатными рецепторами. В 7 интернейронах мы обнаружили значительное увеличение вероятности возникновения антидромного ТД после стимуляции возбуждающих терминалей (p<0,01 по сравнению с контрольными значениями), которое подавлялось при аппликации NBQX, антагониста каинатных рецепторов (100 μМ; p=0,41 по сравнению с контрольными значениями) (Рис. 3.4.4б,в).
Таким образом, мы показали, что активация аксональных каинатных рецепторов может происходить благодаря синаптически высвобождаемому глутамату. Поскольку не существует доказательств наличия аксо-аксональных синапсов в интернейронах поля СА1 гиппокампа, то этот глутамат, по всей видимости, диффундирует от удаленных возбуждающих синапсов и достигает каинатных рецепторов на интернейронах.
3.4.5 Последствия аксональной деполяризации, вызываемой каинатными рецепторами, для гамКергической передачи
В проделанных экспериментах с высокочастотной стимуляцией глутаматергических терминалей нам не удалось показать, что синаптически высвобождаемый глутамат способен вызывать спонтанные токи действия в дистальных участках аксонов. Вероятно, это связано с ограничениями метода исследования гетеросинаптической модуляции in vitro, описанными ранее. Однако, если такой феномен имеет место in vivo, то он может представлять собой абсолютно новый механизм интеграции возбуждающей передачи в гиппокампе. Другими словами, если спонтанные потенциалы действия начинают возникать в аксонах интернейронов, когда внеклеточная концентрация глутамата достигает определенного уровня, то это может лежать в основе гомеостатического механизма, усиливающего торможение в гиппокампе. Однако, эти интернейроны дожны иннервировать пирамидные клетки, тогда как в данной работе мы продемонстрировали, что каинат усиливает торможение и самих интернейронов. Тем не менее, действие каината на спонтанные ТПСТ в пирамидных клетках довольно хорошо исследовано, значительное увеличение в их частоте продемонстрировано несколькими различными лабораториями. Эти данные могут быть объяснены возникновением спонтанных потенциалов действия в дистальном аксоне интернейронов, иннервирующих пирамидные клетки. Другим объяснением является то, что деполяризация в этих клетках возникает за счет активации каинатных рецепторов в соматодендритном компартменте и потенциал действия генерируется в аксонном холмике. Действие каината на спонтанные ТПСТ в интернейронах было изучено гораздо меньше. Незадолго, до публикации наших данных появилась работа Cossart с соавторами, в которой был показан эффект каината на частоту потенциал действия независимых миниатюрных ТПСТ в интернейронах (Cossart et al. 2001b). Это свидетельствует в пользу того, что каинат может, при определенных условиях, влиять непосредственно на высвобождение ГАМК. В наших экспериментах мы не обнаружили этого эффекта на миниатюрные ТПСТ у морских свинок, хотя смогли его воспроизвести при аппликации 1 М (но не 250 нМ) каината на срезах гиппокампа крыс (Рис. 3.4.5). Вероятно, каинат, прежду всего, деполяризует аксоны, но эта деполяризация, при достижении ей определенного уровня, может электротонически распространиться в ГАМКергические терминали, приводя к “прямому” эффекту на высвобождение нейротрансмиттера.
а1
а2
а3
б1
б2
б3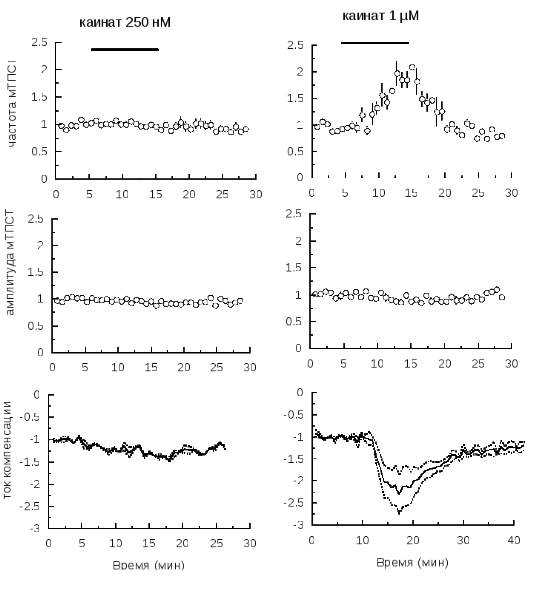
Рис. 3.4.5 Аппликация 1 μМ, но не 250 нМ, каината повышает частоту миниатюрных ТПСТ в интернейронах срезов гиппокампа крыс
а, Аппликация каината 250 нМ не вызывала значительных изменений ни в частоте (а1), ни амплитуде (а2) миниатюрных ТПСТ (спонтанных ТПСТ, регистрируемых в присутствии тетродотоксина) в интернейронах str.radiatum поля СА1 срезов гиппокампа крыс, При этом не происходило изменений в и токе компенсации (а3).б, Аппликация 1 μМ каината приводила к увеличению частоты (б1), но не амплитуды (б2) миниатюрных ТПСТ, а также увеличению тока компенсации (б3).
Для того чтобы определить роль каинатных рецепторов в модуляции ГАМКергической передачи между интернейронами мы исследовали действие каината на спонтанные ТПСТ, многие из которых отражают генерацию спонтанных потенциалов действия в пресинаптических нейронах. Каинат в концентрациях 250 нМ или 1 М значительно увеличивал как частотуспонтанных ТПСТ, регистрируемых в интернейронах, так и их амплитуду.
Существует несколько различных объяснений причинам, вызывающим увеличение одновременно в частоте и амплитуде сТПСТ в интернейронах под действием каината. Например, возможность существования специфической популяции пресинаптических интернейронов, чувствительных к каинату. Другим обяснением может быть облегчение распространения потенциалов действия в местах деления аксонов в пресинаптических интернейронах. Не останавливаясь на неопределенности механизмов, лежащих в основе этого феномена, важно отметить, что увеличение амплитуды сТПСТ под действием каината характерно только для интернейронов и не было показано для пирамидных клеток.
Таким образом, действие каината на нейроны гиппокампа обладает клеткоспецифичностью, подобной той, которая наблюдается в действии агонистов mGluR группы III. Мы показали, что деполяризация пресинаптических аксонов за счет каинатных рецепторов может объяснять увеличение спонтанных ТПСТ. Однако, почему каинат вызывает различные изменения этих токов в интернейронах и пирамидных клетках осталось неясным. Возможно, что различие определяется зависимостью распространения каинатных рецепторов по клеточной мембране в различных пресинаптических интернейронах. Не исключено, что интернейроны, в которых мы регистрировали ТПСТ, представляют собой гомогенную группу из всего разнообразия интернейронов гиппокампа (Freund and Buzsaki 1996; Freund and Gulyas 1997). Таким образом, мы не можем сказать имеют ли эти интернейроны связи с пирамидными клетками или же только с другими интернейронами. С другой стороны, в предыдущих работах не удалось показать определенной корреляции между морфологией интернейронов и действием на них каината (Cossart et al. 1998; Cossart et al. 2001b), что свидетельствует против выборочной модуляции возбудимости нейронов каинатными рецепторами.