

3.2.5 Заключение
Таким образом, в данном разделе работы было показано:
1.В интернейронах str.radiatum, но не пирамидных клетках поля СА1 гиппокампа, при нормальных условиях помимо фазического торможения (ГАМКергических ТПСТ) существует пикротоксин-чувствительный ГАМКергический тонический ток.
2. Увеличение внеклеточной концентрации ГАМК при блокаде ее обратного захвата приводит к возникновению тонического тока как в интернейронах, так и в пирамидных клетках.
3. ГАМКергический тонический ток в интернейронах при нормальных условиях фармакологически отличается от ГАМКергического тонического тока, связанного с повышением внеклеточной концентрации ГАМК.
4. Повышение температуры приводит к увеличению частоты спонтанных ТПСТ (фазических токов). При этом сохраняется клеточная специфичность тонической проводимости для интернейронов, но не пирамидных клеток.
Результаты, полученные в данном разделе работы, являются первым детальным описанием тонического торможения в различных нейронах гиппокампа. Показана клеточная специфичность (интернейрон/пирамидная клетка) и фармакологическое различие типов тонического тока. Эти данные могут быть использованы для селективного изменения возбудимости определенных групп клеток и поиска эффективных антиэпилептических препаратов.
3.3 Модуляция гамКергической передачи в гиппокампе метаботропными рецепторами
Мы произвели регистрацию синаптических токов в интернейронах str.radiatum и пирамидных клетках поля СА1 с помощью пейчкламповских электродов (whole-cell patch pipettes). Приблизительно 25% интернейронов были биполярными клетками, остальные три- и мультиполярными. Мы не обнаружили принципиальных различий в ответах этих типов интернейронов на аппликацию L-AP4 (агонист метаботропных рецепторов группы III) или электрическую стимуляцию пресинаптических терминалей. По этой причине полученные данные были объединены вместе.
3.3.1 L-ap4 подавляет и тормозные, и возбуждающие синаптические токи в интернейронах
Моносинаптические ТПСТ регистрировались при комнатной температуре в ответ на внеклеточную стимуляцию в stratum radiatum в присутствии высоких концентраций антагониста NMDA рецепторов (APV, 50 М) и антагониста AMPA/каинатных рецепторов (NBQX, 10М). Аппликация L-AP4 (агонист метаботропных рецепторов группыIII, 50М) производила обратимое снижение ТПСТ до 4712 % (среднеестандартная ошибка среднего) от контрольных значений (Рис. 3.3.1а; p=0,01; n=5). Неожиданным оказалось то, что эта депрессия была практически идентичной по величине снижению ВПСТ в ответ на аппликацию L-AP4. ВПСТ регистрировались в отдельной группе экспериментов, в которых мы не добавляли NBQX, а заменили его пикротоксином (100М) и бикукуллином (10 µМ), чтобы блокировать ГАМКАрецепторы (Рис. 3.3.1б). В этом случае L-AP4 подавило ВПСТ до 5311 % от контроля (p=0,01, n=5).
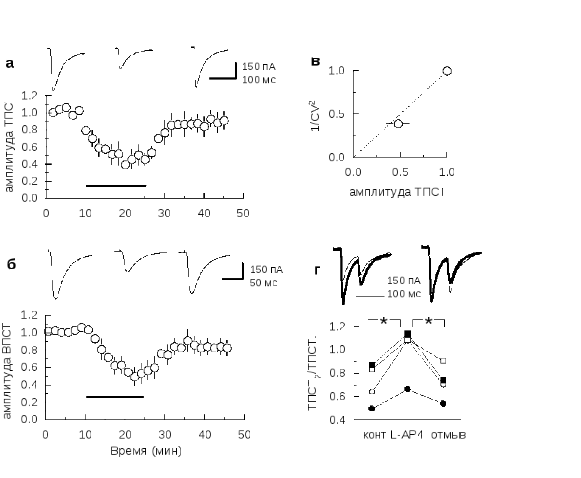
Рис. 3.3.1 L-AP4, агонист метаботропных рецепторов группы III, подавляет моносинаптические ТПСТ и ВПСТ в интернейронах в равной степени
а, Амплитуда ТПСТ (среднее ± С.О.С.), нормированная к средней амплитуде до аппликации L-AP4 (50 μМ) (n=5). Оригинальные записи над графиком получены до, во время и после аппликации L-AP4 с одного нейрона (усреднение по 30 последовательным регистрациям).б, Амплитуда ВПСТ, полученная таким же образом в отдельных сериях экспериментов (n=5).в, Депрессия ТПСТ сопровождалась снижением статистического параметра 1/CV2, что указывает на снижение квантового содержимого (quantal content). Статистический параметр 1/CV2был нормирован к базовым значениям в каждой клетке и усреднен во всех экспериментах. Затем был построен график отношения этого параметра к средней амплитуде ТПСТ, нормированной таким же образом. Диагональная линия показывает траэкторию изменений ожидаемую из распределения Пуассона при снижении квантового параметра высвобожденияm.г, Депрессия, вызываемая L-AP4, сопровождается обратимым изменением коэффициэнта парной депресии, что указывает на пресинаптический сайт действия этого агониста. Оригинальные записи над граффиком получены усреднением 10 последовательных регистраций в одной эксперименте. Жирные линии наложившиеся друг на друга контроль и отмывка, тонкая линия – ТПСТ при аппликации L-AP4. Записи справа теже, что на левой панели, но нормированные к амплитуде первого ТПСТ, чтобы оценить снижение парной депресии. *: p<0,05.
Снижение средней амплитуды ТПСТ сопровождалось снижением статистического параметра 1/CV2(эквивалентного среднее арифметическое2/дисперсия), который изменяется с изменением квантового состава выброса медиатора (Edwards et al. 1989) Было доказано, что его использование позволяет грубо оценить выброс медиатора в гиппокампальных синапсах (Manabe et al. 1993). Полученное изменение 1/CV2в наших экспериментах было пропорционально изменению средней амплитуды (Рис. 3.3.1в). Это свидетельствует в пользу того, что эффект L-AP4 может быть связан с пресинаптическим снижением вероятности выброса нейропередатчика. Для получения дополнительных доказательств пресинаптического эфекта активации метаботропных рецепторов группыIII, мы проверили, какой эффект L-AP4 оказывает на коэффициент парной депрессии ТПСТ. Для этого мы использовали два стимула с интервалом 50 мс. В подтверждение данных, полученных с использованием параметра 1/CV2, коэффициент парной депресии также менялся при аппликацииL-AP4 (Рис. 3.3.1г). Таким образом, мы показали, что ГАМКергические терминали содержат метаботропные рецепторы глутамата, активация которых пресинаптически модулирует выброс ГАМК.
В противоположность значительной депрессии ТПСТ и ВПСТ, регистрируемых в интернейронах, моносинаптические ТПСТ в пирамидных нейронах снижались только до 83 6 % от контроля (Рис. 3.3.2а; p=0,01; n=7). Это снижение было достоверно меньше, чем наблюдаемое в интернейронах (p<0,04; непарный t тест). Кроме того, мы не обнаружили эффекта L-AP4 на полевые ВПСП, записываемые в присутствии пикротоксина и APV вstr.radiatum(Рис. 3.3.2б). Это указывает на то, что и возбуждающая синаптическая передача в пирамидных клетках не чувствительна к активации метаботропных рецепторов группыIII. Эти данные согласуются с результатами предыдущих исследований (Scanziani et al. 1998; Shigemoto et al. 1996),

Рис. 3.3.2 L-AP4 сравнительно неэффективно в снижении ВПСТ и ТПСТ в пирамидных нейронах
а, Нормированная к базовым значениям амплитуда ТПСТ, зарегистрированых в пирамидных нейронах поля СА1, (n=7). Аппликация L-AP4 (50 μМ) приводила к небольшой, но статистически значимой их депрессии. Оргигинальные записи получены до, во время и после аппликации L-AP4 с одного нейрона.б, Наклон полевых ВПСП, полученных в str.radiatum поля СА1 (n=7). Аппликация L-AP4 не приводила к статистически значимому эффекту. Оригинальные записи полученны в одном эксперименте.