

2.8 Статистический анализ
Экспериментальные данные каждой серии (n≥4) были усреднены и представлены в виде средняя ± стандартная ошибка средней. Достоверность различий данных оценивалась с использованием парного или непарного t‑теста и однофакторного дисперсионного анализа (ANOVA). Различия считались достоверными при P<0,05. Аппроксимирующие кривые на графиках рассчитывались по методу наименьших квадратов.
3 Результаты исследованИй и их обсуждение
3.1 Нетипичные фармакологические свойства гамКергических рецепторов в гиппокампальных интернейронах
3.1.1 Различная чувствительность ионотропных гамКергических рецепторов к пикротоксину в интернейронах и пирамидных клетках
Добавление неконкурентного антагониста ионотропных ГАМКергических рецепторов, пикротоксина (PTX), приводило к подавлению ТПСТ как в интернейронах str.radiatum, так и в пирамидных нейронах поля СА1 срезов гиппокампа (Рис.3.1.1 а,б). Однако, чувствительность ТПСТ, регистрируемых в двух популяциях клеток, к этому веществу значительно отличалась. Так использование 100 μМ пикротоксина (концентрация традиционно используемая для подавления ГАМКАрецептор опосредованных ТПСТ) практически полностью подавляло ТПСТ в пирамидных клетках, тогда как в интернейронах все еще регистрировался значительный ток (16 ± 5 % от базовых значений ТПСТ; p<0,01; n=7). Интересно, что аппроксимация по уравнению Хилла кривых концентрация PTX -ингибирование ТПСТ приводила к получению одинакового коэффициента Хилла для обоих типов клеток (0,9), но разному IC50(9 μМ для пирамидных клеток и 26 μМ для интернейронов; Рис.3.1.1в).
Пикротоксин представляет собой рацемическую смесь пикротина и более активного пикротоксинина, который, как предполагается, блокирует ионный канал ГАМКергических рецепторов (Gurley et al. 1995; Newland and Cull-Candy 1992). Хотя известно, что эффект пикротоксина может зависеть от активности рецепторов, нам не удалось обнаружить, что остаточный ТПСТ в интернейронах снижался со временем в присутствии 100 μМ пикротоксина. Таким образом, различная чувствительность ТПСТ
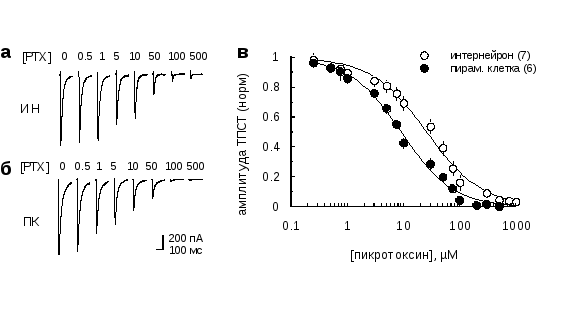
Рис. 3.1.1 ТПСТ в интернейронах менее чувствительны к пикротоксину, чем в пирамидных клетках
а, Оригинальные записи, полученные
с одного интернейрона в присутствии
D-APV (50 μM), NBQX (20 μM), CGP52432 (5M),
каждая усреднена по 10 последовательным
ТПСТ. Пикротоксин подавлял ТПСТ
концентрационно-зависимым образом.
Однако, при концентрации пикротоксина
100 µМ все еще наблюдался значительный
ток. б, оригинальные записи полученные
с одной пирамидной клетки, каждая
усреднена по 10 последовательным ТПСТ.
Пикротоксин, как и в интернейронах,
подавлял ТПСТ в пирамидных клетках
концентрационно-зависимым образом.
Причем, аппликация 100 µМ пикротоксина
полностью подавляла ТПСТ.в,
Зависимости характеризующие доза-эффект
пикротоксина на амплитуду ТПСТ в
интернейронах ( ;
n=7) и пирамидных нейронах (
;
n=7) и пирамидных нейронах ( ;
n=6), построенные в полулогарифмических
координатах. ТПСТ нормированы к амплитуде
базового ТПСТ (в отсутствии пикротоксина).
Линии аппроксимации показывают
теоретические кривые, полученные по
уравнению Хилла. Коэффициент Хила был
одинаковым для обоих типов клеток (0,9),
тогда как IC50была 9 µМ для
интернейронов и 26 µМ для пирамидных
клеток. Разбросы указывают величину
±С.О.С.
;
n=6), построенные в полулогарифмических
координатах. ТПСТ нормированы к амплитуде
базового ТПСТ (в отсутствии пикротоксина).
Линии аппроксимации показывают
теоретические кривые, полученные по
уравнению Хилла. Коэффициент Хила был
одинаковым для обоих типов клеток (0,9),
тогда как IC50была 9 µМ для
интернейронов и 26 µМ для пирамидных
клеток. Разбросы указывают величину
±С.О.С.

в интернейронах и пирамидных нейронах к этому веществу не отражает различную занятость рецепторов эндогенным агонистом (Hajos et al. 2000). С другой стороны, этот результат может быть объяснен какими-то различиями в постсинаптических рецепторах. Таких различий может быть предложено несколько. Во-первых, если ГАМКергические рецепторы гомогенны в обоих типах клеток, то они могут состоять из различных субъединиц (Sperk et al. 1997; Thomson et al. 2000). Во вторых, рецепторы могут быть гетерогенны и, в этом случае, разница в чувствительности ТПСТ в этих типах клеток может объясняться разной пропорцией ГАМКергических рецепторов, слабо чувствительных к пикротоксину. И в третьих, при наличии одинаковых рецепторов в интернейронах и пирамидных клетках их активация может регулироваться разными внутриклеточными механизмами. Для ответа на этот вопрос мы провели разностороннее исследование биофизических и фармакологических свойств ГАМКергических рецепторов в интернейронах и пирамидных клетках.