

2.5.3 Определение биофизических свойств рецепторов с использованием анализа токов, полученных с outside-out пейчей
Нестационарный дисперсионный анализ токов с outside-outпейчей в ответ на быструю аппликацию ГАМК производился, как было описано ранее (Jonas et al. 1993; Perrais and Ropert 1999; Sigworth 1980). Регистрировались как минимум 30 последовательных ответов со стабильной амплитудой (Рис. 2.7). Затем рассчитывался средний ток и дисперсия для каждой точки. Связь между этими параметрами основана на следующих соотношениях:
I(t)=NP(t)i, (1)
где I(t) – средний ток в данный момент времени,N– число каналов открытое при пиковой амплитуде тока,P(t) – вероятность открытого состояния канала в данный момент времени,i– средний ток одиночного канала
σ2ток- σ2шум=NP(t)(1-P(t))i2, (2)
значения параметров те же что и в (1), σ2ток- дисперсия тока, вызванного аппликацией ГАМК, в данной временной точке, σ2шум– дисперсия базового тока до
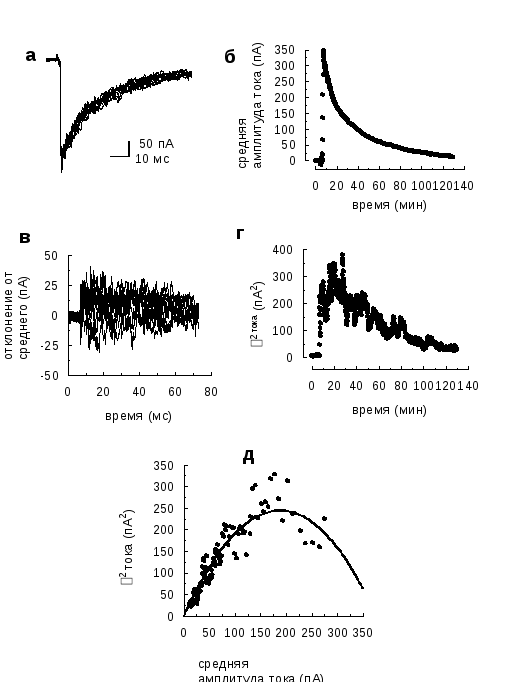
Рис. 2.7 Использование метода нестационарного дисперсионного анализа для определения биофизических параметров ГАМКергических рецепторов
а, Для анализа использовались данные с пейчей, в которых амплитуда ответов на быструю аппликацию была сравнительно стабильной (не наблюдалось тенденции к ее увеличению или уменьшению с течением времени). Ответы оцифровывались с частотой 10 кГц. Полученные токи усреднялись (б) и затем путем вычитания среднего определялась динамика отклонений индивидуальных ответов от среднего (в) После этого рассчитывалась дисперсия для каждой точки ответа, из которой вычиталась дисперсия шума (тока до аппликации агониста;г). Получив эти данные, мы строили зависимость дисперсии от средней амплитуды тока для каждой временной точки (д). Эта зависимость аппроксимировалась по методу наименьших квадратов с использованием уравнения σ2ток- σ2шум= iI-I2/NP, где σ2ток– дисперсия ГАМКергического тока, σ2шум– дисперсия базового тока перед стимуляцией I – средняя амплитуда тока. При этом мы получали параметры: i – средний ток одиночного канала и NPчисло каналов в пейче.
аппликации (вариабельность этого тока связана со спонтанным открыванием-закрыванием каналов).
Из (1) и (2) получается соотношение σ2ток- σ2шум=iI-I2/N, которое связывает амплитуду тока амплитуду тока с дисперсией. Таким образом, зависимость дисперсии от тока аппроксимировалась параболической кривой, полученной по данному уравнению. Из этой аппроксимации мы получали параметрыiиN. После чего рассчитывалась максимальная вероятность открытого состояния канала (Pотк) по уравнению:Pотк=1-(σ2пик- σ2шум)/iIпик, гдеIпики σ2пик– максимальная амплитуда тока и дисперсия в этой точке, соответственно.
2.6 Модели эпилептогенеза in vivo
2.6.1 Электрический киндлинг
Операционная техника
Опыты проводились на 9 крысах-самцах Вистар массой 270-350 граммов. В ходе предварительной операции 5 животным под нембуталовым наркозом (50 мг/кг, в/б) стереотаксически вживлялись 2 электрода в левый гиппокамп: рострально (монополярный электрод; AP=-1,8; ML= 1,5; DV=3,2) и каудально (парный электрод; AP=-3,6; ML=-3; DV=4). Для электрической стимуляции гиппокампа использовался ростральный электрод и один из электродов каудальной пары. Для регистрации послеразрядов (ПР) пирамидных нейронов использовался другой электрод каудальной пары. Индифферентный электрод (для регистрации) находился в назальных частях черепа.
Стимуляция
Внутримозговая электрическая стимуляция животных начиналась через неделю после операции. Был использован протокол (rapid alternate kindling), предложенный ранее Лотманом и Вильямсоном (Lothman and Williamson 1994). Электрической стимуляции подвергались 5 животных. Другие 4 служили в качестве контроля. Частота стимуляции была 50 Гц, длительность 10 секунд. Для стимуляции использовались прямоугольные импульсы тока длительностью 0,2 мсек. и интенсивностью, вызывающей электрические послеразряды (группу популяционных спайков длительностью не менее 10 секунд после окончания стимуляции). Обычно интенсивность тока варьировала в пределах 250-400 μА. В день давалось 12 таких стимулов с интервалом 30 минут. Эти стимулы давались через 1 сутки в течение 8 суток (всего 48 стимулов).
Критерием выработки определенной стадии киндлинга являлась длительность послеразрядов и поведенческие проявления, оцениваемые по шкале Рэйсина (Racine 1972):
1 стадия – "отряхивание мокрой собаки", жевание, подергивание лицевых мышц;
2 стадия – качание головой, судорожные подергивания мышц туловища;
3 стадия – клонус передних конечностей;
4 стадия - клонус передних конечностей и поднимание на задние лапы;
5 стадия - клонус передних конечностей, поднимание на задние лапы и потеря равновесия.
Срезы гиппокампа между ростральным и каудальным электродами выделялись через 24 часа после того, как животные достигали 4-5 стадий киндлинга (соответствующие электрографические и поведенческие реакции на 3 последовательных стимуляции).