

Часть III. Патофизиология органов и систем
эффекторов (аденилатциклаза, гуанилатцикла-за, фосфолипаза С) и накоплении циклических аденозинмонофосфата (цАМФ) и гуанозинмоно-фосфата или диацилглицерола и инозинтрифос-фата. Вторичные посредники, в свою очередь, запускают последующую цепь процессов, важнейшими звеньями которых являются активация протеинкиназ и фосфорилирование белковых субстратов. По такому механизму, в частности, катехоламины, регулируют интенсивность гликогенолиза. Специфичность ответа клетки на тот или иной гормон определяется специфичностью рецептора, который связывается только со своим гормоном, а также природой специфических для клетки протеинкиназ и белковых субстратов.
Другая группа гормонов проникает в клетку, где оказывает свое действие. Эту группу можно обозначить как группу гормонов «непосредственного» действия. Сюда входят андрогены, эстрогены, прогестины, кортикостероиды. Главным в действии стероидных гормонов является активация или торможение того или иного гена, что сопровождается усилением или угнетением образования соответствующего фермента. Однако ряд эффектов осуществляется другими путями, не связанными с влиянием на активность генов. В механизме доставки стероида к генетическому локусу можно выделить три звена. Первое звено - связывание поступившего в клетку гормона с белком, находящимся в цитоплазме и выполняющим роль специфического рецептора цля данного гормона. Второе звено - модификация комплекса «стероид + рецепторный белок». Эта модификация дает возможность осуществления следующего - третьего звена. Третье звено - проникновение стероида в комплексе с рецепторами в ядро клетки и избирательное соединение со специфическим участком хроматина.
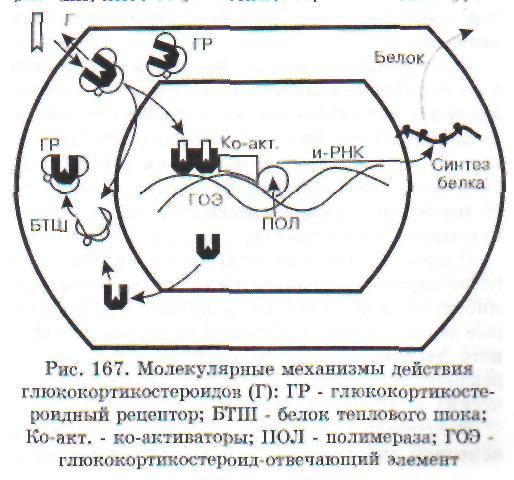
Общий механизм влияния гормонов «непосредственного» действия можно проиллюстрировать на примере глюкокортикоидов (рис. 167). Гормон свободно проникает в клетку и связывается со специфическими рецепторными белками цитоплазмы - глюкокортикоидными рецепторами (ГР). Очевидно, связывается неметаболи-зированный гормон, поскольку из стероидно-белкового комплекса удается выделить глюкокор-тикоид как таковой. Об этом свидетельствует и тот факт, что метаболиты кортизола не вызывают эффектов кортизола и конкурентно не угнетают его действия. Рецепторные белки обладают вы-
соким сродством к стероиду, выраженной специфичностью и малой емкостью. Поэтому данный вид связывания называют специфическим. В зависимости от вида клеток количество рецепторов колеблется от 3000 до 5000 на одну клетку. Сравнение различных тканей одного вида животных показало, что связывание глюкокор-тикоида различно в разных тканях. Так, растворимая фракция клеток тимуса связывала в 3 раза больше триамсинолона, чем такие же фракции из коры головного мозга и тестикул.
Глюкокортикоидные рецепторы относятся к суперсемейству стероид-ядерных регуляторных протеинов, которые функционируют как лиганд-активируемые факторы транскрипции. В цитоплазме ГР в несвязанном с гормоном состоянии представляют собой гетерогенные комплексы, состоящие из собственно рецептора и связанных с ним по крайней мере четырех белков теплового шока (БТШ). Роль последних заключается в поддержании конформации ГР в состоянии, подходящем для связывания гормона и предупреждения транслокации несвязанного с гормоном ГР в ядро. После связывания ГР с гормоном он освобождается из комплекса с белками теплового шока и перемещается в ядро. Здесь глюкокор-тикоид-рецепторный комплекс превращается в димер и связывается в регуляторной части соответствующего гена с определенным участком ДНК, называемым гормон-отвечающим элементом (ГОЭ). ГР-димер регулирует транскрипцию этого гена, вызывая либо активацию транскрипции, либо ее угнетение. При снижении уров-
631
ня гормона и диссоциации гормонально-рецеп-торного комплекса в ядре ГР освобождается и возвращается обратно в цитоплазму, где снова образует комплекс с белками теплового шока.
Препараты глюкокортикоидных гормонов обычно применяют для подавления воспаления при многих заболеваниях (аутоиммунные процессы, бронхиальная астма и др.). Механизмы их антивоспалительного действия многообразны и реализуются через изменение регуляции активности многих генов, кодирующих образование провоспалительных цитокинов, ферментов и других продуктов, участвующих в развитии воспаления. Так, глюкокортикоиды:
усиливают экспрессию генов, кодирующих образование ферментов, оказывающих угнетающее влияние на развитие воспаления (липокор-тин-1, нейтральная эндопептидаза в эпителиальных клетках слизистой дыхательных путей разрушают тахикинины, лейкоцитарный секрети-руемый ингибитор протеазы в слизистой дыхательных путей и др.);
угнетают экспрессию генов, кодирующих образование провоспалительных цитокинов (ин-терлейкины-1 -6, гранулоцитарно-макрофагаль-ный колониестимулирующий фактор, фактор некроза опухоли и др.);
угнетают экспрессию генов, кодирующих образование энзимов, способствующих развитию воспаления (синтетаза оксида азота, индуцибель-ная изоформа циклоксигеназы-2);
угнетают экспрессию генов, кодирующих образование молекул адгезии (ICAM-I) и рецепторов для провоспалительных медиаторов (для вещества Р).
Существуют и другие механизмы действия этих гормонов, которые изучены меньше и объединяются под названием посттранскрипционных. Сюда входят все звенья, начиная со стабилизации и транспорта образовавшихся и-РНК и до окончания синтеза белка. Во многих точках этого многозвеньевого процесса глюкокортикоиды оказывают свое влияние.
Одним из важных механизмов в действии глюкокортикоидов является так называемое пер-миссивное действие. Оно означает, что некоторые метаболические эффекты гормонов дистантного действия, о которых упоминалось выше, реализуются только в присутствии физиологических концентраций глюкокортикоидов.
Все гормоны, циркулирующие в организме, в конечном счете метаболизируются и выводятся
632
из него. В основном метаболизм гормонов происходит в печени. Однако ряд гормонов метабо-лизируется и в других тканях.
В организме для каждого гормона существует равновесие между его секрецией, связыванием белками, действием в тканях-мишенях и метаболизмом в тканях. В поддержании такого равновесия большую роль играет механизм обратной связи. Нарушение любого из внежелези-стых компонентов этого равновесия может приводить к таким изменениям, которые будут клинически проявляться как нарушение функции соответствующей железы.
Нарушение связывания гормонов белками
Связывание кортикостероидов белками плазмы крови, будучи одним из эволюционно-выра-ботавшихся физиологических механизмов приспособления, очевидно, при определенных условиях может нарушаться, и тогда это нарушение связывания может стать патогенетическим фактором либо сниженной, либо повышенной физиологической активности кортикостероидных гормонов. Клинические наблюдения указывают на такую возможность. Так, например, при синдроме Иценко - Кушинга выявляются случаи, сопровождаемые снижением связывания корти-зола белками плазмы крови, что приводит к увеличению свободной фракции кортизола. При снижении способности белков плазмы крови связывать кортизол обнаруживали также признаки диабета или преддиабета, нарушения менструального цикла, гипертензию и др. Нарушение связывания тиреоидных гормонов может приводить к таким изменениям, которые определяются как гипо- или гипертиреоз. Усиление связывания инсулина может приводить к картине инсулиновой недостаточности.
Блокада циркулирующего гормона
Этот вид изменения активности гормона касается полипептидных гормонов. Механизмы такого изменения различны, но общим результатом будут такие изменения в организме, которые соответствуют картине гипофункции соответствующей железы.
Возможны следующие механизмы инактивации:
а) инактивация гормона в связи с образованием аутоантител к тому или иному гормону.