

Механизм сокращения мышцы
Сокращение мышц – это результат укорочения каждого ее саркомера, которое происходит путем вдвигания актиновых нитей между миозиновыми нитями.
Различают L (изотропный) диск, образованный нитями миозина. Повторяющиеся участки миофибрилл между двумя L-дисками называют саркомерами, их длина 2,5 мкм. При сокращении каждый саркомер укорачивается на 20-50 %, причем это сокращение происходит за счет параллельного скольжения тонких (актин) и толстых (миозин) нитей относительно друг друга.
Скольжение нитей актина и миозина друг относительно друга сопровождается поочередным образованием и разрушением поперечных мостиков между этими нитями за счет конформационных изменений в нитях миозина.
Химическим пусковым механизмом к началу сокращения служит выход катионов кальция Са++ из цистерн. В результате ионы кальция специфически стимулируют Са2+-зависимую АТФ- азную активность актомиозина, в возбужденной мышце происходит распад АТФ на АДФ и неорганический фосфат с освобождением энергии, которая используется для сокращения саркомера. Сигналом для выхода ионов Са++ из цистерн в саркоплазму служит электрический импульс, приходящий из двигательного нерва через моторную пластинку. Такой импульс приводит к разрядке трансмембранного потенциала мышечных волокон. При этом резко увеличивается проницаемость мембран (Т-система) и ионы Ca2+ выходят из цистерн, создавая условия реакции сокращения. С прекращением электрического импульса ионы кальция под действием кальциевого насоса вновь поглощаются Т-системой.
Источником энергии для мышечного сокращения является АТФ. Для сохранения работоспособности должен происходить постоянный ресинтез АТФ. Одним из основных источников ресинтеза АТФ является гликоген мышц. В процессе гликолиза гликоген превращается в молочную кислоту, а энергия, освобождающаяся при этом, идет на синтез АТФ. В мышцах кроме АТФ имеется и другой макроэрг – креатинфосфат, концентрация которого в 4-5 раз больше, чем АТФ.
Продолжительная работа мышц приводит к истощению запасов креатинфосфата и в этих условиях ресинтез АТФ из АДФ может идти за счет гликолиза и тканевого дыхания. При высокой мышечной активности для получения энергии используется главным образом глюкоза. Переход мышц из покоя к работе ведет к увеличению потребления кислорода в ней в 20 раз и более. Одновременно уменьшается отношение АТФ:АДФ, что приводит к активации фосфофруктокиназы и ферментов цикла трикарбоновых кислот. АДФ может также вступать в реакцию перефосфорилирования друг с другом:
АДФ + АДФ => АТФ + АМФ
Это является дополнительным механизмом ресинтеза АТФ.
В период покоя в мышцах происходит накопление креатинфосфата:
АТФ + креатин => креатинфосфат + АДФ
Повышенная нагрузка на мышцы (быстрый бег, поднятие больших грузов) сопровождается учащением дыхания вследствие усиления гликолиза, что приводит к накоплению молочной кислоты в ткани. Молочная кислота используется затем для ресинтеза гликогена в печени и мышечной ткани (гликонеогенез).
Азотистые экстрактивные вещества мышц
Это - вещества, извлекаемые из мышц при настаивании с водой, содержание их по азоту составляет 300-500 мг %. К ним относятся: креатин, карнозин, ансерин, карнитин, глутаминовая кислота, глутамин, аминокислоты, мочевина, пептиды, АТФ.
Среди небелковых азотистых веществ самое высокое содержание креатина – 60 % всего небелкового азота мышц.
Креатин образуется из глицина, аргинина, метионина. Его образование начинается в печени. Мышцы адсорбируют креатин из крови и удерживают в значительных количествах. Более половины креатина находится в виде креатинфосфата, образующегося в результате реакции трансфосфорилирования между креатином и АТФ. Креатинфосфорная кислота является дополнительным резервом энергии. Она запасается в покоящейся мышце и интенсивно используется для синтеза АТФ в работающей мышце. Передавая неорганический фосфат на АДФ, креатин необратимо превращается в креатинин, удаляемый из организма с мочой.
Синтез креатина происходит из аргинина, глицина и метионина:
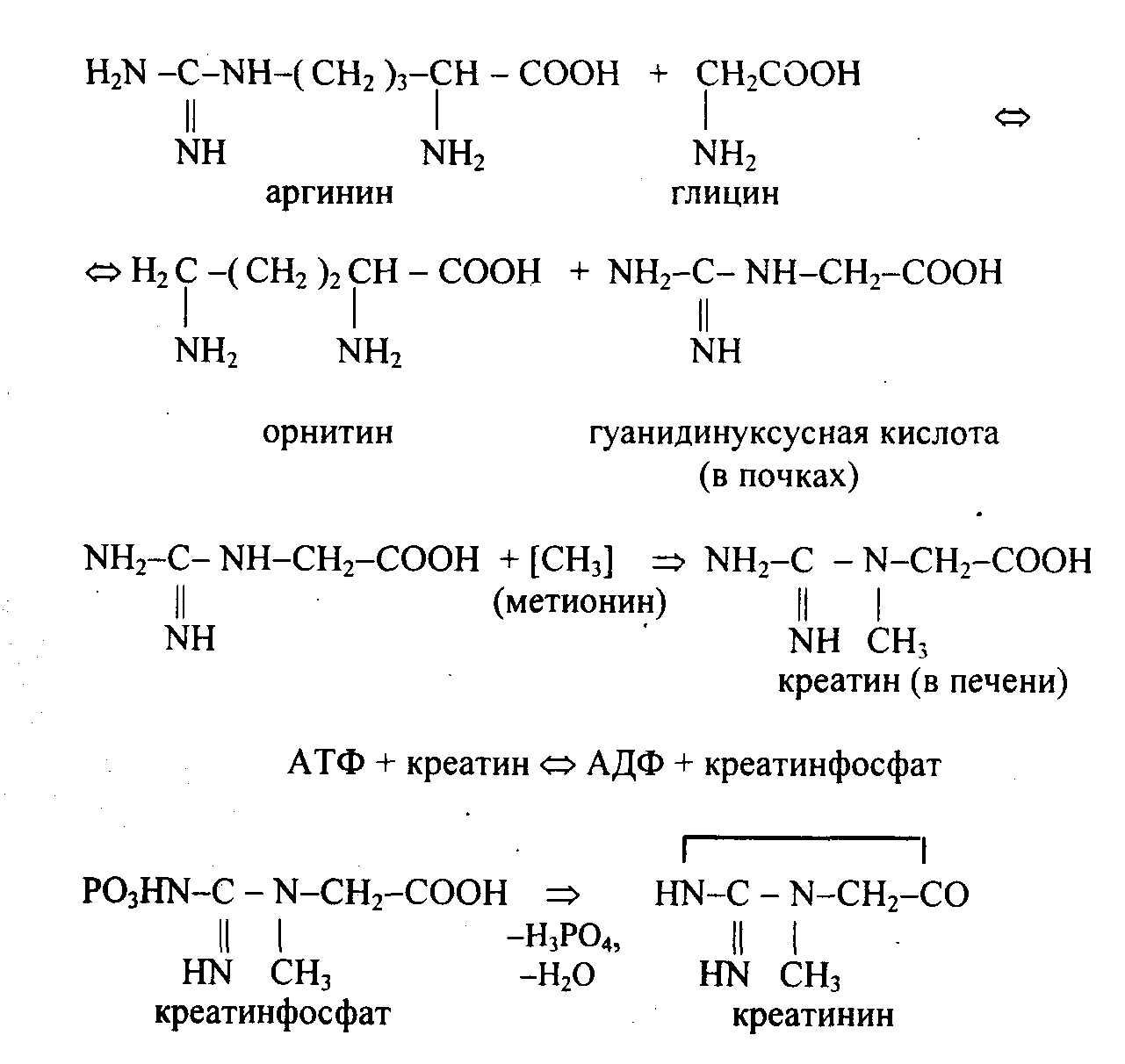
При болезнях мышц увеличивается концентрация креатина в крови и выделение его с мочой. Суточное выделение креатина в норме величина постоянная. Содержание его в крови в норме 1-2 мг/дл. При болезнях почек с нарушением фильтрации выделение креатина уменьшается, а его концентрация в крови увеличивается.
К специфическим азотистым экстрактивным веществам скелетных мышц относятся карнозин, ансерин, карнитин.
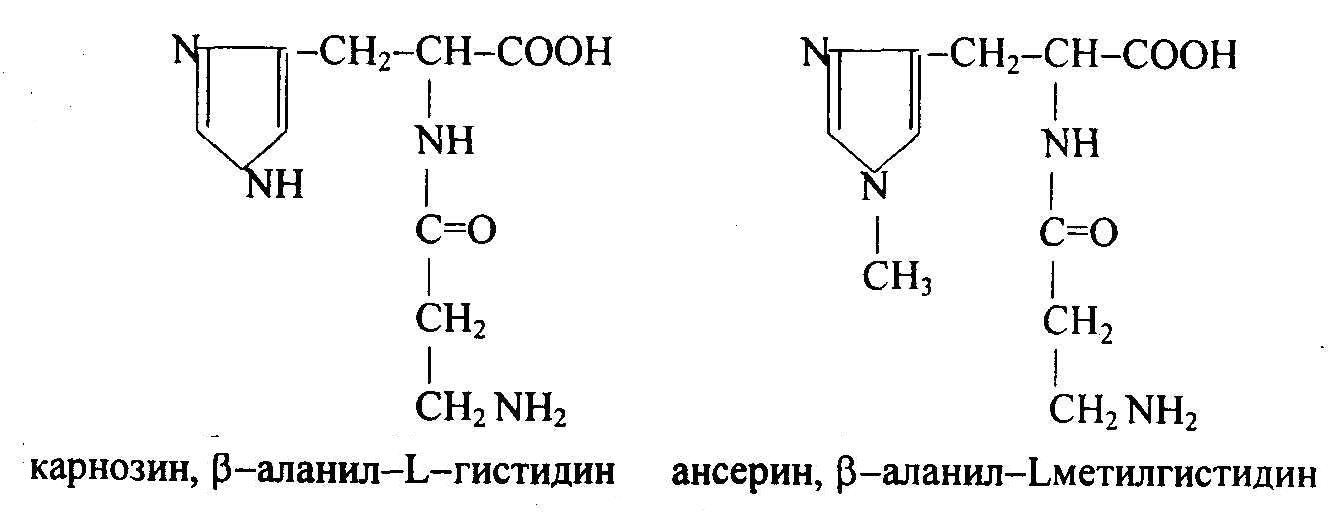
Дипептиды карнозин и ансерин были открыты А.С. Гуливичем и его сотрудниками в 1900 г; эти пептиды в больших количествах содержатся в белых мышцах (0,2-0,3 %). Е.С. Северин показал, что они увеличивают амплитуду мышечного сокращения, создают буферную емкость быстрых мышц, являются антиоксидантами, комплексонами металлов, защитными веществами, предотвращающими модификацию белков. Карнозин оказывает защитное действие на ряд ферментов и ферментных комплексов, а также на ионные насосы, является гидрофильным антиоксидантом, понижает в клетках уровень свободных радикалов.
Карнитин участвует в транспорте жирных кислот через мембрану митохондрий, тем самым, ускоряя использование жирных кислот для синтеза АТФ в работающей мышце.
Глутаминовая кислота и глутамин (150-200 мг %) играют важную роль в обезвреживании аммиака, образующегося во время мышечной работы.