

Особенности переваривания белков у жвачных животных
Как известно, у жвачных животных желудок 4-х камерный, состоит из рубца, сетки, книжки, сычуга. Сычуг является собственным желудком, где вырабатывается сок, содержащий HCl, реннин, пепсин. Рубец, сетка, книжка – в них происходит основное превращение белков и других азотистых веществ корма под действием микрофлоры и простейших. Рубцовая микрофлора имеет сбалансированный характер. Общее количество бактерий в рубце составляет 109-1010 клеток в 1 г содержимого, или в рубцовом содержимом находится 1,5-2,0 кг бактерий. Это огромное количество. Под действием бактериальных ферментов растительные и животные белки в рубце расщепляются до пептидов, аминокислот и свободного аммиака. Одновременно идет синтез бактериального белка за счет размножения микробов.
Кроме бактерий в рубце значительное место занимают простейшие, их количество в 1 мл составляет 105-106. На долю инфузории приходится 1/20 массы рубцовой жидкости у крупного рогатого скота, а их азот занимает 10-12 % общего азота кормовой смеси преджелудков. Инфузории размножаются быстро, за сутки меняется 4-5 поколений и ежесуточное их переваривание в сычуге и кишечнике достигает 70 % всех простейших рубца.
Простейшие синтезируют незаменимые аминокислоты, обеспечивающие животное полноценным белком. Это имеет существенное значение для животного. Простейшие содержат больше незаменимых аминокислот, особенно лизина, чем бактерии. Таким образом, в организме жвачных бактерии – главный поставщик ферментов, гидролизирующих белки до аминокислот.
Бактерии и инфузории систематически поступают в сычуг и перевариваются. Часть аминокислот под действием ферментов бактерий подвергаются гидролитическому дезаминированию:
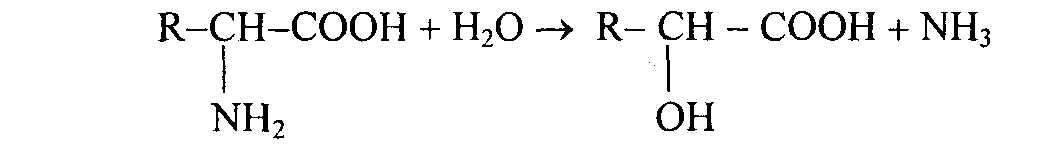
Или происходит восстановительное дезаминирование с участием НАДН2:
![]()
В результате процессов дезаминирования аминокислот в рубце накапливается большое количество аммиака. Оптимальным для бактерий и инфузорий является значение рН 5,5-7,0. После дезаминирования кислотные остатки превращаются в CO2 и летучие жирные кислоты. Микрофлора рубца способна расщеплять не только белковые молекулы, но и небелковые азотистые вещества. Известно, что вместе с растительными кормами до 10-30 % азота может быть, в виде небелкового азота (аминокислоты, амиды и т.д.). Эти продукты гидролизуются до аммиака и жирных кислот. Таким же процессам гидролиза подвергаются пуриновые и пиримидиновые основания: аденин, гуанин, ксантин, мочевая кислота и гуанидин, которые расщепляются с образованием NH3, CO2 и уксусной кислоты.
Мочевина (диамид угольной кислоты) может попадать в рубец с кормом или слюной (куда попадает из крови в результате обезвреживания аммиака в печени в орнитиновом цикле), а также в виде подкормки (карбамид-мочевина). При недостаточности азотистых веществ в корме мочевина начинает усиленно поступать в рубцовое содержимое путем выделения стенкой рубца из крови.
В рубце под влиянием бактериальных ферментов уреазы мочевина гидролизуется с образованием аммиака, CO2, H2O:
![]()
Аммиак частично всасывается в кровь и превращается в печени в мочевину, далее выделяется почками. Частично используется бактериями для синтеза аминокислот:
Бактерии могут синтезировать все аминокислоты заменимые и незаменимые, о чем уже указывалось выше.
Для практики животноводства важно использование карбамида для пополнения азота рациона жвачных, при этом следует учесть, что гидролиз мочевины идет в 4 раза быстрее, чем усвоение бактериями освободившегося аммиака, поэтому в корм добавляют мочевину малыми порциями, в брикетированном виде, в виде добавок к силосу (добавляют при силосовании) и т.д. За сутки коровы могут использовать 60-150 г мочевины, овцы 10-15 г. Это позволяет заменить 20-30 % перевариваемого протеина корма.
Распад белков в тканях и его биологическое значение
Аминокислоты всасываются в кровь, доставляются в печень, где частично дезаминируются, декарбоксилируются или подвергаются трансаминированию. Кроме того, происходит постоянно обновление белков собственного тела – распад (в лизосомах) и синтез de novo. Обновление аминокислот в белках ткани идет очень интенсивно. Так, белки печени обновляются наполовину за 8-12 суток, плазмы крови – за 18-45 суток. У крупного рогатого скота при выращивании на мясо за сутки синтезируется 120-200 г белка, у лактирующей коровы с молоком выделяется 600-1200 г новых белков. Распад тканевых белков – аутолиз происходит под действием ферментов – тканевых протеаз – катепсинов.
Третьим источником свободных аминокислот (1-ый из кишечника, 2-ой - аутолиз) в клетках организма является их синтез. В растениях синтезируется очень большой набор аминокислот (свыше 20), а в животном организме синтезируются только заменимые аминокислоты путем восстановительного аминирования кетокислот и трансаминирования.
Восстановительное аминирование кетокислот является обратным процессом окислительного дезаминирования аминокислот (глутаминовая, аспарагиновая и др.). Ресинтез происходит в 2 этапа:

Таким образом, в первую фазу реакции из кетокислоты и аммиака образуются иминокислоты, во вторую – иминокислота восстанавливается за счет водорода восстановленной формы НАД или НАДФ, то есть НАД•H2, НАДФ•Н2 – в аминокислоту. Этот путь синтеза аминокислот в организме животных ограничен, он ярче выражен у растений и микробов (бактерий).
Наиболее выраженный путь биосинтеза аминокислот в организме – путь переаминирования (трансаминирования). Он открыт в 1937 году Браунштейном A.E. и Крицманом М.Г. Было установлено, что из глутаминовой и пировиноградной кислот могут образоваться α-кетоглутаровая кислота и аланин без промежуточного выделения аммиака.

Эту реакцию называют трансаминированием, при этом происходит перенос аминогруппы с аминокислоты на кетокислоту. Донором аминогруппы является аминокислота, акцептором – кетокислота. Все природные аминокислоты подвержены ферментативному переаминированию. Наиболее активно эта реакция происходит между глутаминовой кислотой и щавелевоуксусной.
Между аспарагиновой кислотой и α-кетоглутаровой (в печени и мышечной ткани) реакция происходит с участием трансфераз (трансаминаз); коферментом является фосфо- пиридоксаль (витамин B6).
Аминогруппа через основание Шиффа переходит на фосфопиридоксаль, в результате синтезируется фоофопиридоксамин и соответствующая кетокислота. Фосфопиридоксамин реагирует с новой кетокислотой, образуя новую аминокислоту с освобождением фосфопирид оксаля. Процесс образования промежуточного продукта можно представить следующим образом:
Процесс образования промежуточного продукта можно представить следующим образом:

Переаминирование играет очень важную роль при синтезе в тканях необходимых аминокислот.
Таким образом, фонд свободных аминокислот клеток формируется за счет:
1) поступления из органов пищеварения;
2) распада белков;
3) синтеза заменимых аминокислот в реакциях трансаминирования, восстановительного аминирования кетокислот.