

Окислительное расщепление аминокислот
Большую часть энергии организм получает в результате окисления углеводов и нейтральных жиров (до 90 %). Остальную часть ~ 10% за счет окисления аминокислот. Аминокислоты, прежде всего, используются для синтеза белка. Окисление их происходит:
1) если аминокислоты, образующиеся при обновлении белков не используются для синтеза новых белков;
2) если в организм поступает избыток белка;
3) в период голодания или при сахарном диабете, когда нет углеводов или их усвоение нарушено, в качестве источника энергии используются аминокислоты.
Во всех этих ситуациях аминокислоты теряют свои аминогруппы и превращаются в соответствующие α-кетокислоты, которые затем окисляются до СО2 и H2O. Частично это окисление идет через цикл трикарбоновых кислот. В результате дезаминирования и окисления образуются пировиноградная кислота, ацетил-КоА, ацетоацетил-КоА, α-кетоглутаровая кислота, сукцинил-КоА, фумаровая кислота. Некоторые аминокислоты могут превращаться в глюкозу, а другие – в кетоновые тела.
Пути обезвреживания аммиака в тканях животных
Аммиак токсичен, и накопление его в организме может привести к смерти. Существуют следующие пути обезвреживания аммиака:
1. Синтез аммонийных солей.
2. Синтез амидов дикарбоновых аминокислот.
3. Синтез мочевины.
Синтез аммонийных солей происходит ограниченно в почках, это как дополнительное защитное приспособление организма при ацидозах. Аммиак и кетокислоты частично используются для ресинтеза аминокислот и для синтеза других азотистых веществ. Кроме того, в тканях почек аммиак участвует в процессе обезвреживания оргинических и неорганических кислот, образуя с ними нейтральные и кислые соли:
R – COOH + NH3 → R – COONH4;
H2SO4 + 2 NH3 → (NH4)2SO4;
H3PO4 + NH3 → NH4H2PO4
Этим путем организм защищается от потери с мочой при выведениикислот знпачительного количества катионов (Na, K, отчасти Са, Mg), что могло бы привести к резкому снижению щелочного резерва крови. Количество аммонийных солей, выводимых с мочой, заметно повышается при ацидозе, так как аммиак используется для нейтрализации кислоты. Одним из путей связывания и обезвреживания аммиака является использование его для образования амидной связи глутамина и аспарагина. При этом из глутаминовой кислоты под действием фермента глутаминсинтетазы синтезируется глутамин, из аспарагиновой кислоты при участии аспарагинсинтетазы – аспарагин:

Этим путем происходит устранение аммиака во многих органах (мозг, сетчатка, почки, печень, мышцы). Амиды глутаминовой и аспарагиновой кислот могут образоваться и тогда, когда эти аминокислоты находятся в структуре белка, то есть акцептором аммиака может быть не только свободная аминокислота, но и белки, в состав которых они входят. Аспарагин и глутамин доставляются в печень и используются в синтезе мочевины. Аммиак переносится в печень и с помощью аланина (глюкозо-аланиновый цикл). Этот цикл обеспечивает перенос аминогрупп из скелетных мышц в печень, где они превращаются в мочевину, а работающие мышцы получают глюкозу. В печени глюкоза синтезируется из углеродного скелета аланина. В работающей мышце из α-кетоглутаровой кислоты образуется глутаминовая кислота, которая затем передает аминную группу - NH2 пировиноградной кислоте, в результате синтезируется аланин – нейтральная аминокислота. Схематически указанный цикл выглядит следующим образом:
Глутаминовая кислота + пировиноградная кислота ↔
↔ α-кетоглутаровая кислота + аланин
 Рис.
10.1. Глюкозо-аланиновый цикл.
Рис.
10.1. Глюкозо-аланиновый цикл.
Этот цикл выполняет две функции: 1) переносит аминогруппы из скелетных мышц в печень, где они превращаются в мочевину;
2) обеспечивает работающие мышцы глюкозой, поступающей с кровью из печени, где для ее образования используется углеродный скелет аланина.
Образование мочевины – основной путь обезвреживания аммиака. Этот процесс изучали в лаборатории И.П.Павлова. Показано, что мочевина синтезируется в печени из аммиака, CO2 и воды.
Мочевина выводится с мочой в качестве главного конечного продукта белкового, соответственно аминокислотного обмена. На долю мочевины приходится до 80-85% всего азота мочи. Главным местом синтеза мочевины в организме является печень. Сейчас доказано, что синтез мочевины происходит в несколько этапов.
1 стадия – образование карбамоилфосфата происходит в митохондриях под действием фермента карбомоилфосфат-синтетазы:

На следующей стадии с участием орнитина синтезируется цитруллин:
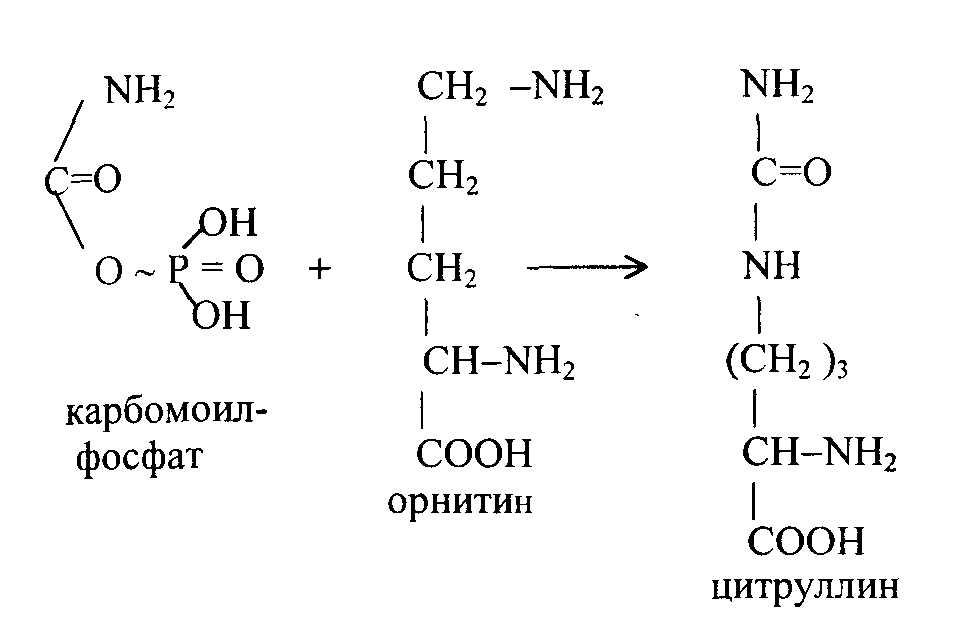
Цитруллин переходит из митохондрий в цитозоль клеток печени. После этого в цикл вводится вторая аминогруппа в форме аспарагиновой кислоты. Происходит конденсация молекул цитруллина и аспарагиновой кислоты с образованием аргинин-янтарной кислоты.

Цитруллин аспарагиновая аргинин-янтарная
кислота кислота
Аргинин-янтарная кислота расщепляется на аргинин и фумаровую кислоты.

Под действием аргиназы аргинин гидролизуется, образуется мочевина и орнитин. В дальнейшем орнитин поступает в митохондрии и может включиться в новый цикл обезвреживания аммиака, а мочевина выделяется с мочой.
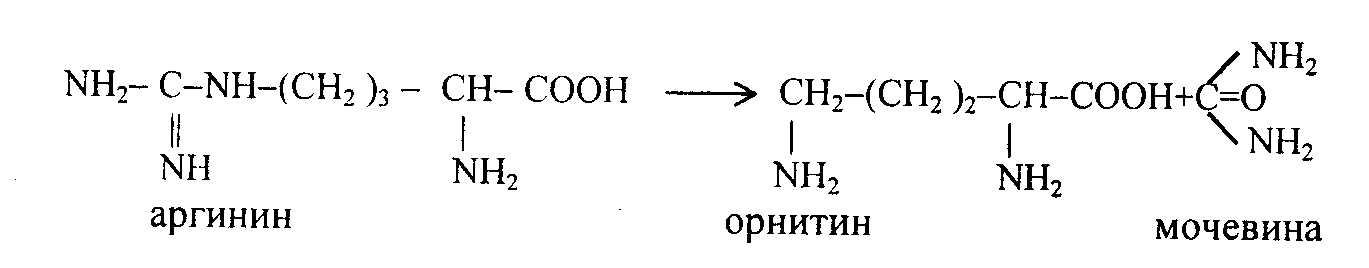
Таким образом, в синтезе одной молекулы мочевины нейтрализуется две молекулы NH3 и CO2 (HCO3), что также имеет значение в поддержании рН. Для синтеза одной молекулы мочевины расходуется 3 молекулы АТФ, в том числе две при синтезе карбомоилфосфата, одна для образования аргинин-янтарной кислоты; фумаровая кислота может превращаться в яблочную и щавелевоуксусную кислоты (цикл Кребса), а последняя в результате трансаминирования или восстановительного аминирования может превратиться в аспарагиновую кислоту. Некоторая часть азота аминокислот выделяется из организма в виде креатинина, который образуется из креатина и креатинфосфата.
Из всего азота мочи на долю мочевины приходится до 80-90%, аммонийных солей – 6 %. При избыточном кормлении белком доля азота мочевины возрастает, а при недостаточном белковом кормлении снижается до 60 %.
У птиц и рептилий – нейтрализация аммиака происходит путем образования мочевой кислоты. Птичий помет на птицефабриках - это источник азотсодержащего удобрения (мочевая кислота).