

Двойной липидный слой мембран
Молекулы фосфолипидов и гликолипидов амфифильны, то есть углеводородные радикалы жирных кислот и сфингозина являются гидрофобными, а другая часть молекулы, образованная из углеводов, остатка фосфорной кислоты с присоединенным к нему холином, серином, этаноламином – гидрофильна. В результате этого в водной среде гидрофобные участки молекулы фосфолипидов вытесняются из водной среды и взаимодействуют между собой, а гидрофильные участки контактируют с водой, в результате образуется двойной липидный слой клеточных мембран (рис.9.1.). Этот двойной слой мембраны пронизан белковыми молекулами – микротрубочками. На наружной стороне мембраны прикреплены олигосахариды. Количество белка и углеводов в различных мембранах неодинаково. Белки мембран могут выполнять структурные функции, могут быть ферментами, осуществлять трансмембранный перенос питательных веществ, могут выполнять различные регуляторные функции. Мембраны всегда существуют в виде замкнутых структур (см. рис.9.1). Липидный бислой обладает способностью к самосборке. Эту способность мембран используют для создания искусственных липидных пузырьков – липосом.
Липосомы широко применяются как капсулы для доставки различных лекарственных веществ, антигенов, ферментов в различные органы и ткани, так как липидные капсулы способны проникать через клеточные мембраны. Это позволяет направлять лекарственные вещества точно по адресу в пораженный орган.

Рис.9.1. Схема клеточной мембраны из двойного липидного слоя. Гидрофобные участки молекулы липидов притягиваются между собой; гидрофильные участки молекулы находятся с наружной стороны. Молекулы белков пронизывают липидный бислой.
Обмен липидов
В организме нейтральные жиры находятся в 2-х формах: запасного жира и протоплазматического жира.
В состав протоплазматического жира входят фосфолипиды и липопротеиды. Они участвуют в формировании структурных компонентов клеток. Мембраны клеток, митохондрий и микросом состоят из липопротеидов и регулируют проницаемость отдельных веществ. Количество протоплазматического жира стабильно, и не изменяется в зависимости от голодания или ожирения.
Запасной (резервный) жир – в его состав входят триацилглицерины жирных кислот – находится в подкожной жировой клетчатке и в жировых депо внутренних органов.
Функции резервного жира заключаются в том, что это -запасной источник энергии, доступной для использования в период голодания; это – изоляционный материал от холода, от механических травм.
Важно также, что липиды, распадаясь, выделяют не только энергию, но и значительное количество воды:
При окислении 1 грамма белка выделяется – 0,4 г; углеводов – 0,5 г; липидов – 1 г воды. Это свойство липидов имеет большое значение для животных, обитающих в условиях пустыни (верблюды).
Переваривание липидов в желудочно-кишечном тракте
В полости рта липиды подвергаются лишь механической обработке. В желудке имеется небольшое количество липазы, которая гидролизует жиры. Малая активность липазы желудочного сока связана с кислой реакцией содержимого желудка. Кроме того, липаза может влиять только на эмульгированные жиры, в желудке отсутствуют условия для образования эмульсии жира. Только у детей и у моногастричных животных липаза желудочного сока играет важную роль в переваривании липидов.
Кишечник является основным местом переваривания липидов. В двенадцатиперстной кишке на липиды воздействует желчь печени и сок поджелудочной железы, одновременно происходит нейтрализация кишечного содержимого (химуса). Происходит эмульгирование жиров под действием желчных кислот. В состав желчи входят: холевая кислота, дезоксихолевая (3,12 дигидроксихолановая), хенодезоксихолевая (3,7 дигидроксихолановая) кислоты, натриевые соли парных желчных кислот: гликохолевая, гликодезоксихолевая, таурохолевая, тауродезоксихолевая. Они состоят из двух компонентов: холевой и дезоксихолевой кислот, а также глицина и таурина.
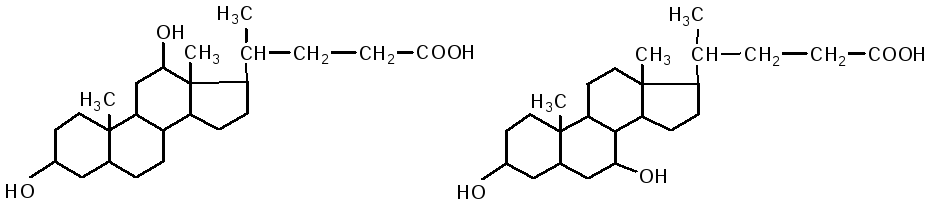
дезоксихолевая кислота хенодезоксихолевая кислота
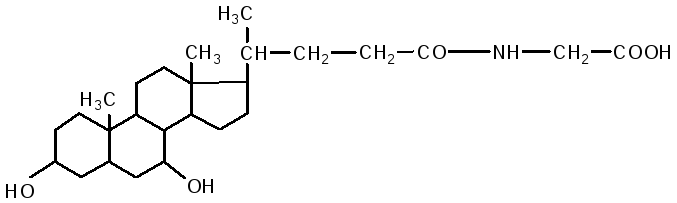
гликохолевая кислота
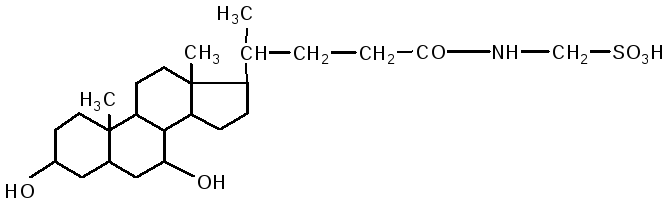
таурохолевая кислота
Соли желчных кислот хорошо эмульгируют жиры. При этом увеличивается площадь соприкосновения ферментов с жирами и увеличивается действие фермента. Недостаточность синтеза желчных кислот или задержка поступления нарушает эффективность действия ферментов. Жиры, как правило, всасываются после гидролиза, но часть тонко эмульгированных жиров всасывается через стенку кишечника и переходит в лимфу без гидролиза.
Эстеразы разрывают в жирах эфирную связь между, спиртовой группой и карбоксильной группой карбоновых кислот и неорганических кислот (липаза, фосфатазы).
Под действием липазы жиры гидролизуются на глицерин и высшие жирные кислоты. Активность липазы возрастает под действием желчи, т.е. желчь непосредственно активирует липазу. Кроме того, активность липазы увеличивают ионы Са++ вследствие того, что ионы Са++ образуют нерастворимые соли (мыла) с освободившимися жирными кислотами и предотвращают их подавляющее влияние на активность липазы.
Под действием липазы в начале гидролизуются эфирные связи у α и α1 (боковых) углеродных атомов глицерина, затем у β-углеродного атома:
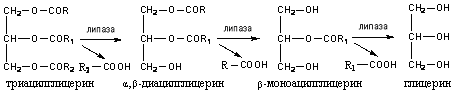
Под действием липазы до 40% триацилглицеридов расщепляются до глицерина и жирных кислот, 50-55% гидролизуется до 2-моноацилглицеринов и 3-10% не гидролизуется и всасываются в виде триацилглицеринов.
Стериды корма расщепляются ферментом холестеролэстеразой до холестерина и высших жирных кислот. Фосфатиды гидролизуются под влиянием фосфолипаз А, A2, С и D. Каждый фермент действует на определенную сложноэфирную связь липида. Точки приложения фосфолипаз представлены на схеме:
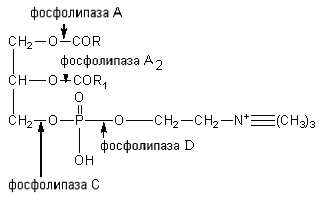
Фосфолипазы поджелудочной железы, тканевые фосфолипазы вырабатываются в виде проферментов и активируются трипсином. Фосфолипаза A2 змеиных ядов катализирует отщепление ненасыщенной жирной кислоты в положении 2 фосфоглицеридов. При этом образуются лизолецитины с гемолитическим действием.

фосфотидилхолин лизолецитин
Поэтому при попадании этого яда в кровь происходит сильный гемолиз.. В кишечнике эта опасность устраняется действием фосфолипазы A1, быстро инактивирующей лизофосфатид в результате отщепления от него остатка насыщенной жирной кислоты с превращением его в неактивный глицерофосфохолин.
Лизолецитины в малых концентрациях стимулируют дифференцировку лимфоидных клеток, активность протеинкиназы С, усиливают клеточную пролиферацию.
Коламинфосфатиды и серинфосфатиды расщепляются фосфолипазой А до лизоколаминфосфатидов, лизосеринфосфатидов, которые далее расщепляются фосфолипазой A2. Фосфолипазы С и D гидролизуют связи холина; коламина и серина с фосфорной кислотой и остатка фосфорной кислоты с глицерином.
Всасывание липидов происходит в тонком отделе кишечника. Жирные кислоты с длиной цепи менее 10 углеродных атомов всасываются в неэтерифицированной форме. Для всасывания необходимо присутствие эмульгирующих веществ – желчных кислот и желчи.
Ресинтез жира, характерного для данного организма, происходит в кишечной стенке. Концентрация липидов в крови в течение 3-5 часов после приема корма высокая. Хиломикроны – мелкие частицы жира, образующиеся после всасывания в кишечной стенке, представляют собой липопротеиды, окруженные фосфолипидами и белковой оболочкой, внутри содержат молекулы жира и желчных кислот. Они поступают в печень, где липиды подвергаются промежуточному обмену, а желчные кислоты проходят в желчный пузырь и далее обратно в кишечник (см. рис.9.3 на стр.192). В результате такого кругооборота теряется малое количество желчных кислот. Считают, что молекула желчной кислоты в сутки совершает 4 кругооборота.