

Особенности обмена отдельных аминокислот
Глицин – легко синтезируется в организме животных, лишь для птиц может быть лимитирующей аминокислотой.
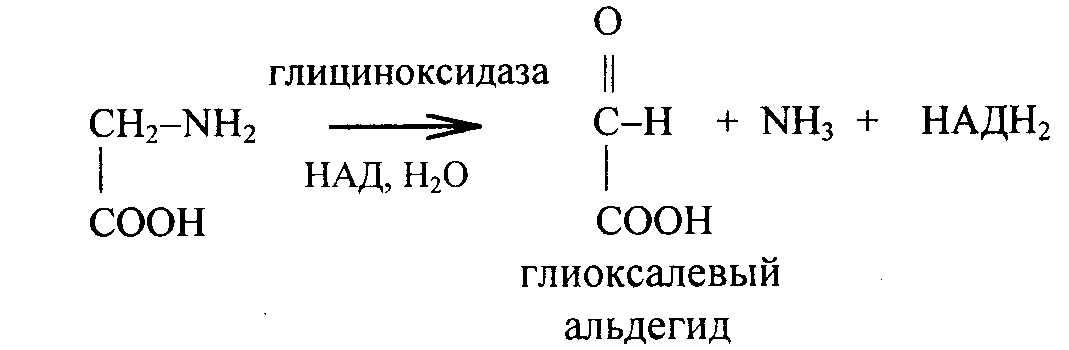
Дезаминируется в тканях под влиянием глициноксидазы с образованием глиоксалевого альдегида. При этом НАД восстанавливается в НАДН2, который в дыхательной цепи митохондрий дает три молекулы АТФ. Глицин используется для синтеза парных желчных кислот, глутатиона, креатина, серина, коламина, пуринов, порфиринов. Идет на обезвреживание бензойной и фенилуксусной кислот.
Серин – при дезаминировании его образуется пировиноградная кислота и аммиак.
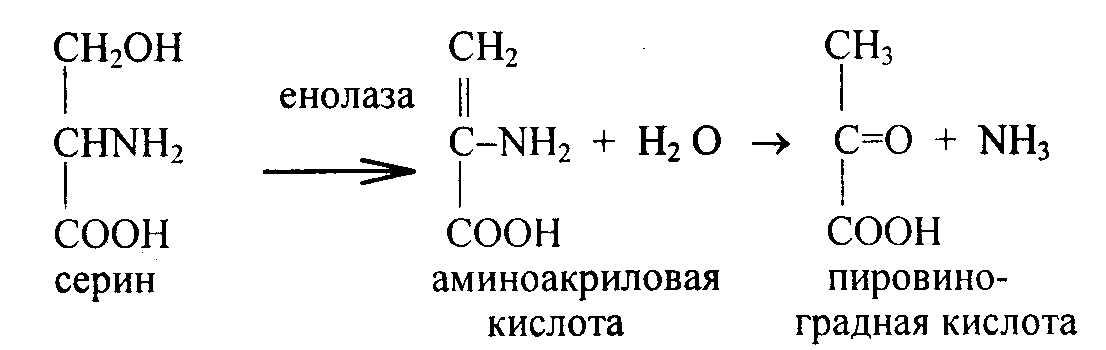
Серин входит в состав серинсодержащих фосфолипидов, является исходным продуктом образования этаноламина и холина, цистеина.
Общую схему катаболизма и глюконеогенез можно представить в следующем виде (рис.10.2., по Николаеву А.Я.):
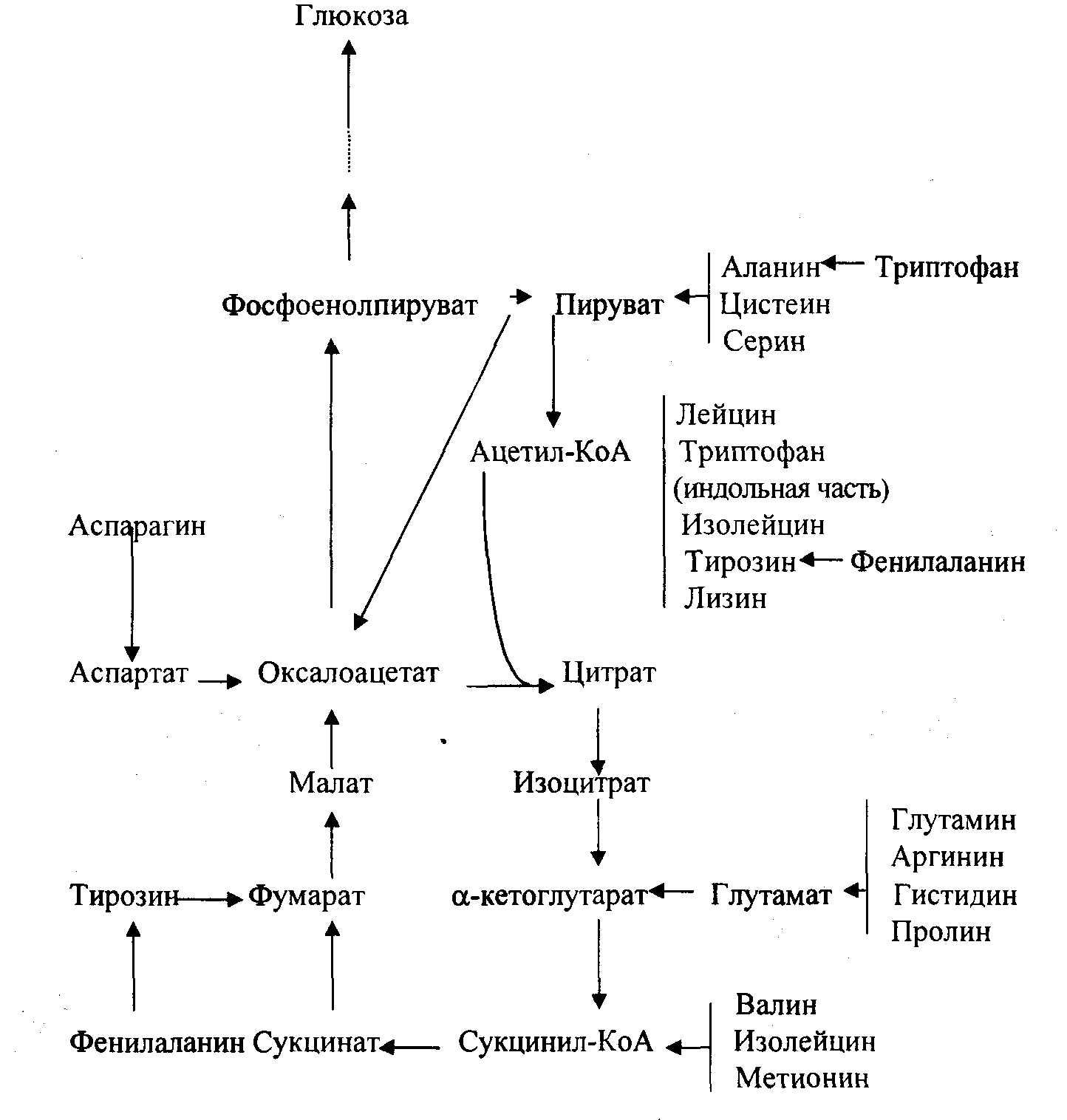
Рис. 10.2. Введение аминокислот в общий путь катаболизма и глюконеогенез.
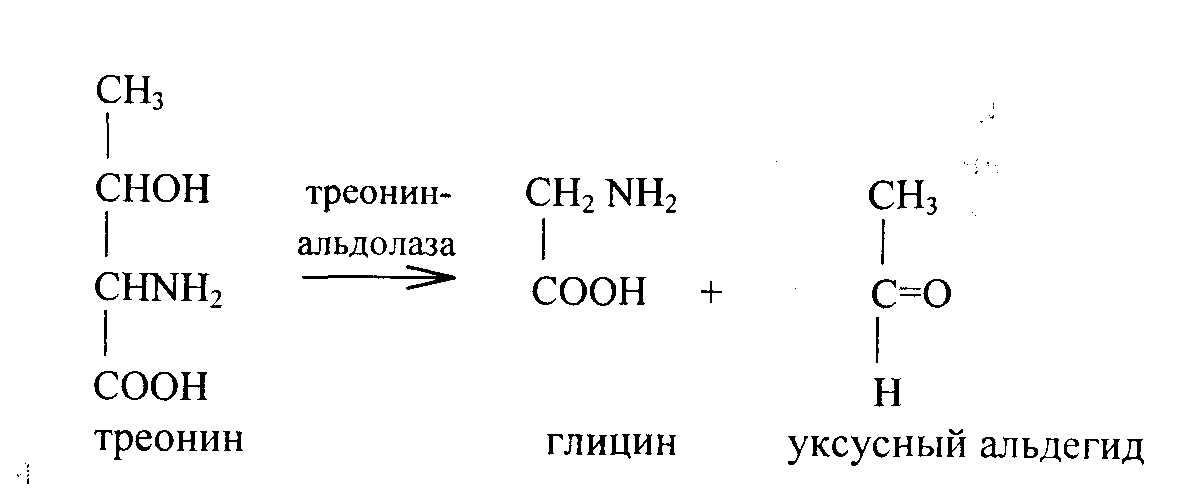
Треонин - незаменимая аминокислота для всех видов животных. Под действием альдолазы превращается в глицин и уксусный альдегид.
Цистеин и цистин. К недостатку серосодержащих аминокислот чувствителен крупный рогатый скот и овцы. Цистеин и цистин легко превращаются друг в друга путем окислительно-восстановительных реакций:
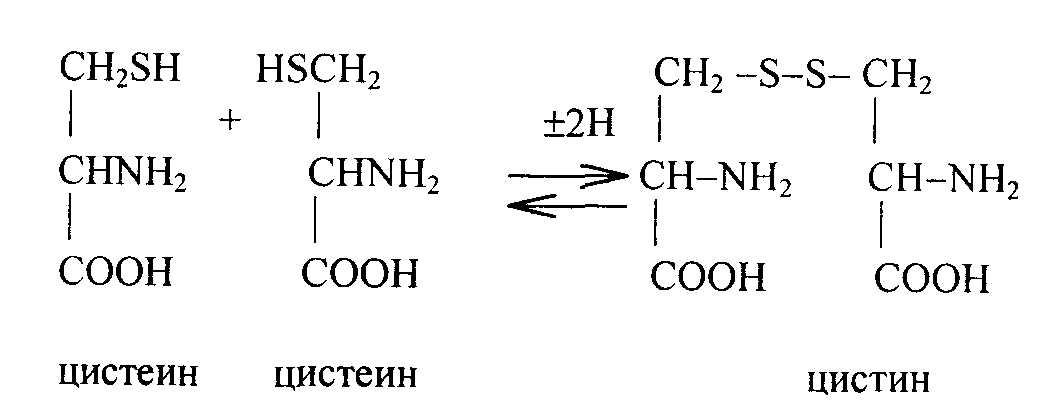
Наличие –SH, -S-S- группировок определяет высокую реактивность ферментов и гормонов. Часть цистеина превращается в таурин, который используется в синтезе парных желчных кислот.
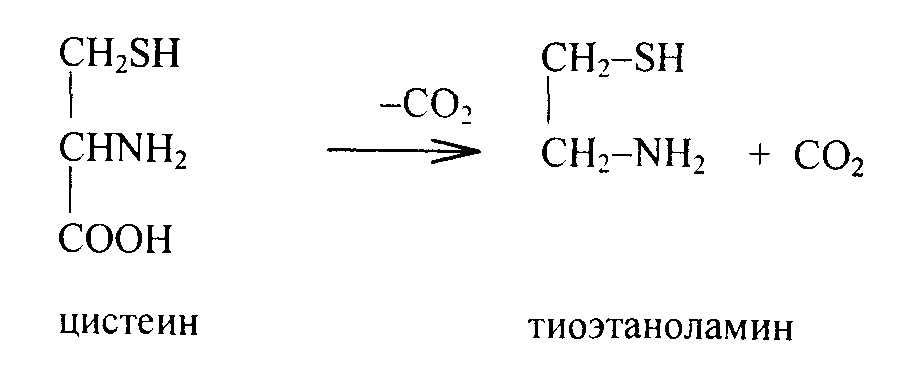
При декарбоксилировании цистеина образцется тиоэтаноламин – кофактор фермента активирования кислот HS-KoA.
Цистеин входит в состав глутатиона – трипептида, широко представленного в эритроцитах, печени, который может быть в восстановленной (HS-глутатион) и окисленной (-S-S-) формах. Глутатион является кофактором дегидрогеназы 3-фосфоглицеринового альдегида и глиоксилазы.
Метионин – незаменимая аминокислота, принимает участие в синтезе цистеина. Метионин имеет CH3 метильную группу активную в трансметилировании. Это универсальный донор метальных групп (для этаноламина, карнозина, гуанидинуксусной кислоты, норадреналина, пиримидиновых оснований).
Аспарагиновая и глутаминовая кислоты. Их много в белках растений. Играют роль в трансаминировании, дезаминировании других аминокислот. Синтезируются из кетокислот. Глутамин используется в синтезе пуриновых оснований мононуклеотидов. При декарбоксилировании аспарагиновой кислоты может образоваться β и α-аланин:
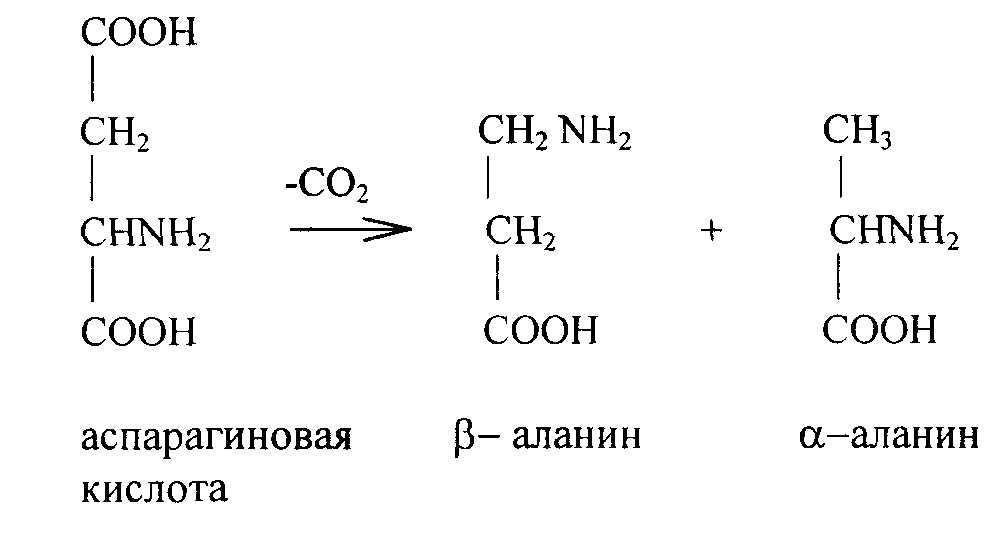
β-аланин идет на синтез пантотеновой кислоты. При декарбоксилировании глутаминовой кислоты образуется у-аминомасляная кислота.
Лизин – незаменимая аминокислота. Биологический распад лизина проходит по сложному пути с образованием α-аминоадипиновой, α- кетоадипиновой и глутаровой кислот.
Фенилаланин и тирозин являются субстратами для синтеза тироксина, адреналина, норадреналина. Валин, лейцин, изолейцин – превращения их направлены на синтез жирных кислот и кетоновых тел. Остальные аминокислоты и два амида могут служить субстратами для синтеза глюкозы и гликогена. Глюконеогенез из аминокислот (синтез глюкозы) происходит интенсивно из гликогенных аминокислот при преимущественно белковом кормлении животных или голодании. При голодании используются белки собственных тканей.
Катаболизм лейцина и лизина не включает стадии образования пировиноградной кислоты.