

Метаболизм ацетил-коэнзима а
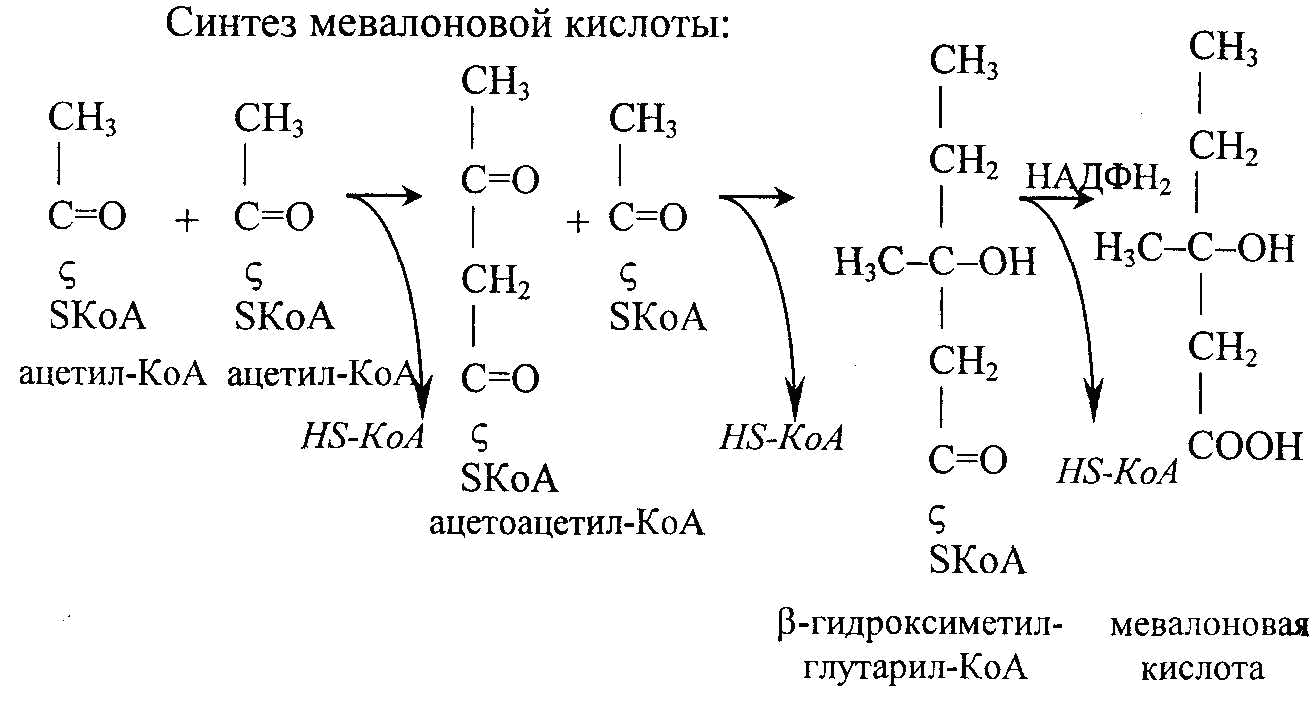
Ацетил-КоА может окисляться в цикле трикарбоновых кислот. При этом обеспечивается 12 молекул АТФ на каждую молекулу ацетилкоэнзима А. Ацетил-КоА может использоваться для синтеза жирных кислот, при этом требуется достаточное количество энергии (АТФ). При недостатке энергии, при нарушении обмена углеводов и липидов синтез холестерина через мевалоновую кислоту нарушается и происходит образование кетоновых тел с их накоплением в тканях и выделением с мочой (кетозы). Мевалоновоя кислота является предшественником холестерина. Однако реакции биосинтеза холестерина почти полностью совпадают с реакциями биосинтеза кетоновых тел. При нормальной энергетической обеспеченности ацетоацетил-КоА полностью превращается в печени в мевалоновую кислоту, затем в холестерин. При уменьшении углеводов, сахарном диабете, нарушении обмена липидов происходит увеличение кетоновых тел, развиваются кетозы.
Пути образования кетоновых тел
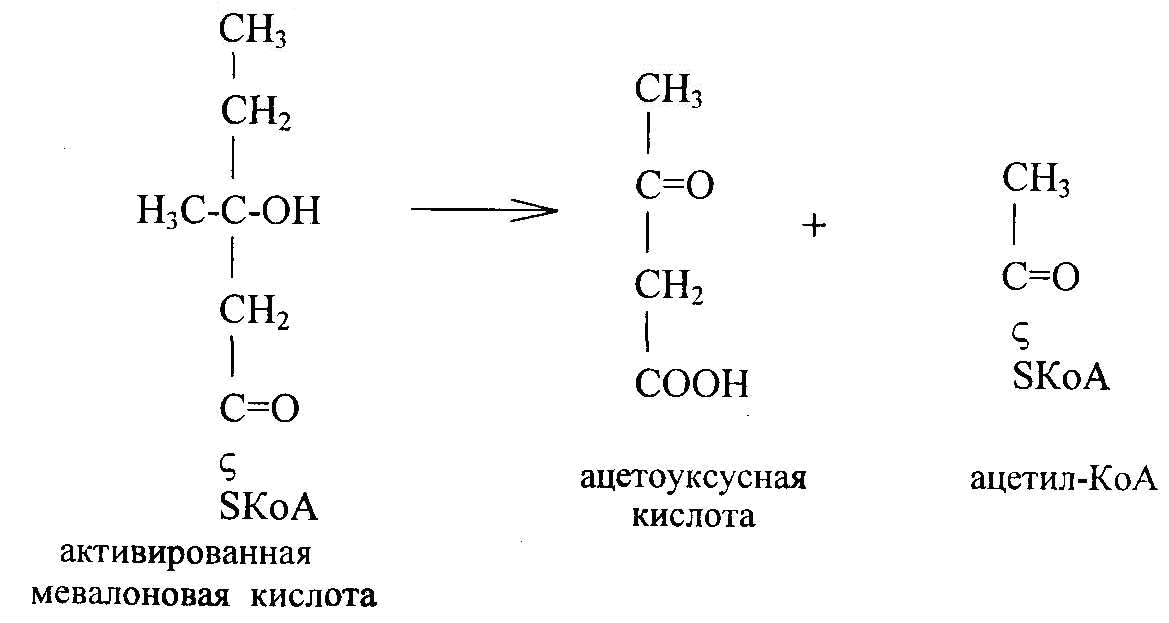
1. Образование кетоновых тел происходит при углеводном голодании, уменьшении энергетических источников для синтеза жира, холестерина из активированной уксусной кислоты. При этом активированная мевалоновая кислота распадается на ацетоуксусную кислоту и ацетил-КоА:
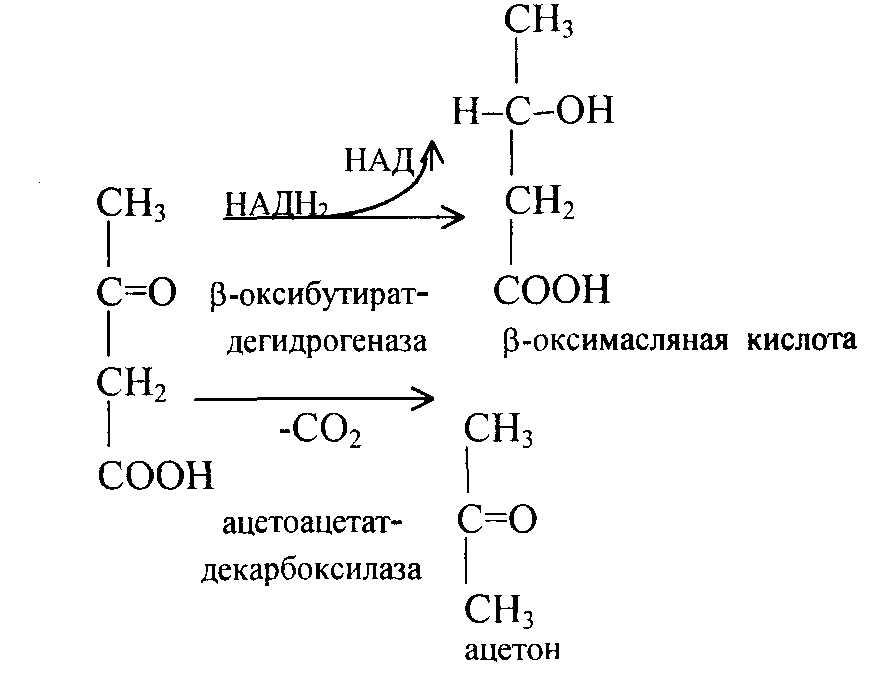
Ацетоуксусная кислота частично восстанавливается до (β-оксимасляной кислоты, а частично декарбоксилируется, образуя ацетон:
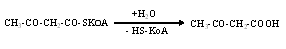
II. Если CH3-CO-KoA образуется много и недостаточно энергии для синтеза жира, то образуется активированная ацетоуксусная кислота:

Затем образуется свободная ацетоуксусная кислота:
Образующаяся ацетоуксусная кислота может превращаться в β-оксимасляную кислоту или ацетон, как указано выше. Кетоновые тела синтезируются в печени и поступают в кровь. В нормальных условиях β-оксимасляная кислота, ацетоуксусная кислота окисляются в тканях мышц и являются источником энергии. Однако при нарушениях обмена (сахарный диабет), когда использование углеводов затруднено и усиливается окисление липидов, при длительном недостатке углеводов, происходит чрезмерный биосинтез кетоновых тел с образованием ацетона. При этом происходит кетонемия (ацидоз) и нарушение нормального функционирования организма.
Окисление ненасыщенных жирных кислот происходит после их превращения в соответствующую насыщенную жирную кислоту. Затем они окисляются по β-углеродному атому.
Окисление жирных кислот с нечетным числом углеродных атомов происходит так же, как и при окислении жирных кислот с четным числом атомов углерода. Отличие состоит в том, что в конечном итоге образуется пропионил-КоА, который вначале подвергается карбоксилированию, в результате образуется активированная янтарная кислота (сукцинил-КоА). Сукцинил-КоА окисляется в цикле трикарбоновых кислот.
Биосинтез липидов
Способность животных запасать полисахариды довольно ограничена; поэтому глюкоза, поступающая в организм в значительных количествах, используется для синтеза жирных кислот и глицерина. Важным процессом является также биосинтез холестерина, так как он является исходным продуктом для образования многочисленных биологически активных стероидов.
Синтез высших жирных кислот происходит из ацетил-KoA в цитозоле. Фермент ацетил-КоА-карбоксилаза, содержащий биотин, катализирует АТФ-зависимый синтез малонил-КоА из ацетил-КоА и СО2:
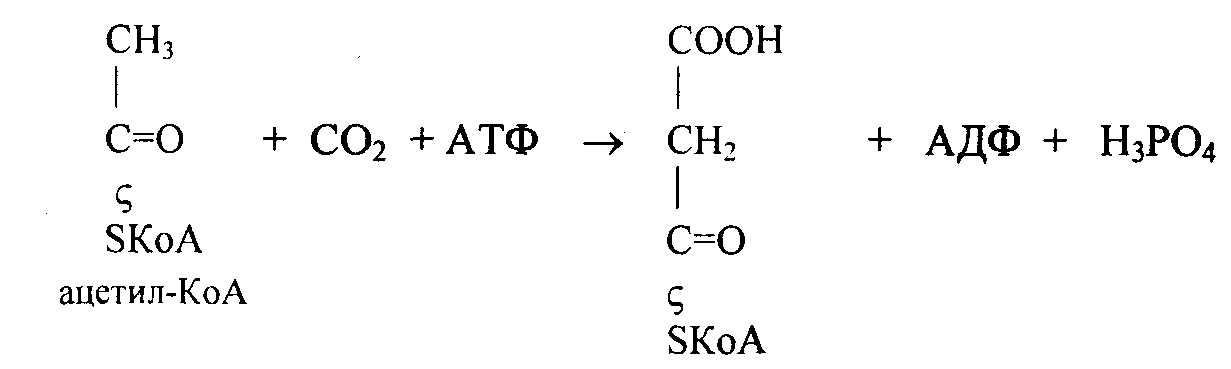
![]()
При участии ацилпереносящего белка (АПБ) Мм 10 Да происходит синтез ацетил-SАПБ и малонил-SАПБ:
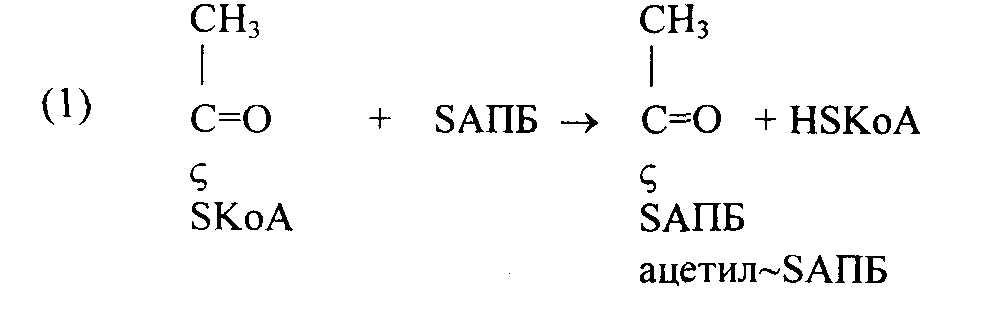
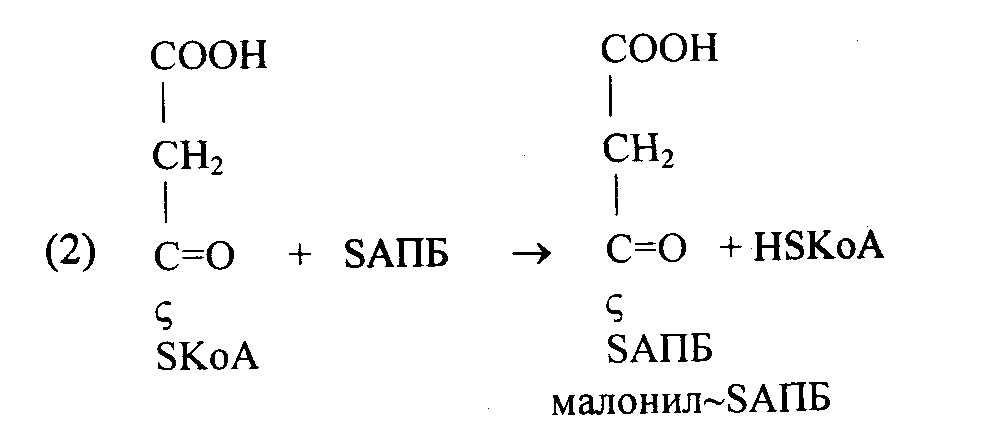
На следующем этапе ацетил-SАПБ и малонил-SАПБ взаимодействуют с образованием ацето-ацетил- SAПБ:
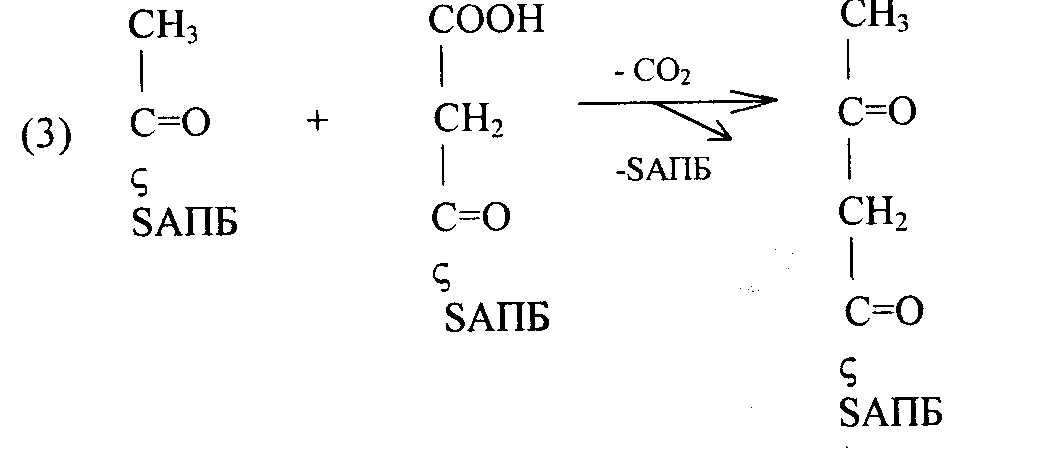
ацетоацетил-SАПБ
После этого ацетоацетил-SАПБ восстанавливается по β-углеродному атому с участием НАДФН2, в результате образуется β-гидроксибутирил-SAПБ. Затем происходит присоединение активированной уксусной кислоты и молонила к ацетилпереносящему белку (АПБ):
β-Гидроксибутирил~SАПБ под действием фермента β-гидрокси-АПБ-дегидратазы теряет молекулу воды (5) и превращается в кротонил-SАПБ; последний с участием НАДФН2 восстанавливается в бутирил-SAПБ. Далее цикл реакции повторяется.
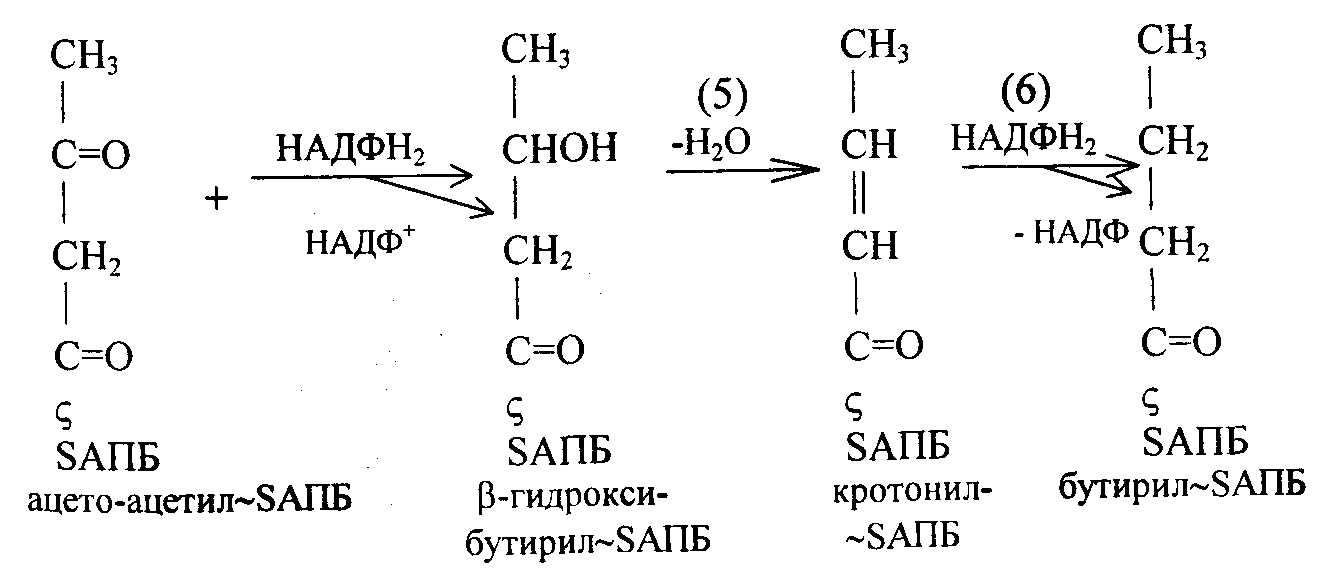
Если идет синтез пальмитиновой кислоты (C16), то к бутирил-SAПБ присоединяется последовательно еще 7 раз малонил~SАПБ, при этом отщепляется дистальная карбонильная группа малонил~SАПБ в виде CO2. Суммарный результат этих реакций следующий:
а цетил-КоА
+ 7 малонил-КоА + 14 НАДФН2
цетил-КоА
+ 7 малонил-КоА + 14 НАДФН2
п альмитиновая
кислота + 7CO2
+ 8 HSKoA
+ 14 НАДФ + 7 H2O
альмитиновая
кислота + 7CO2
+ 8 HSKoA
+ 14 НАДФ + 7 H2O
В этом процессе 7 молекул CO2 образуется за счет свободных карбоксильных групп 7 молекул малонил-КоА. Из 16 углеродных атомов пальмитиновой кислоты 2 атома образуется за счет ацетил-КоА, а остальные за счет малонил-КоА.
В ходе синтеза 7 карбоксильных групп – С=О- восстанавливаются из семи групп -CH2 -, на это расходуется 14 НАДФН2: за счет 7 из них образуются водородные атомы групп -CH2 -, а за счет остальных семи – кислород карбонильных групп превращается в воду.
Биосинтез фосфатидных кислот и триглицеридов происходит в печени и жировой ткани при наличии глицерина и жирных кислот. Глицерин образуется при гидролизе липидов, затем под влиянием фермента глицерокиназы с участием АТФ синтезируется глицерофосфат:

Глицерофосфат может образоваться также из диоксиацетонфосфата, возникшего при окислении глюкозы путем восстановления НАДН2.

фосфодиоксиацетон глицерофосфат
Глицерофосфат далее используется для биосинтеза фосфатидной кислоты. Реакция катализируется фосфатидсинтетазой:
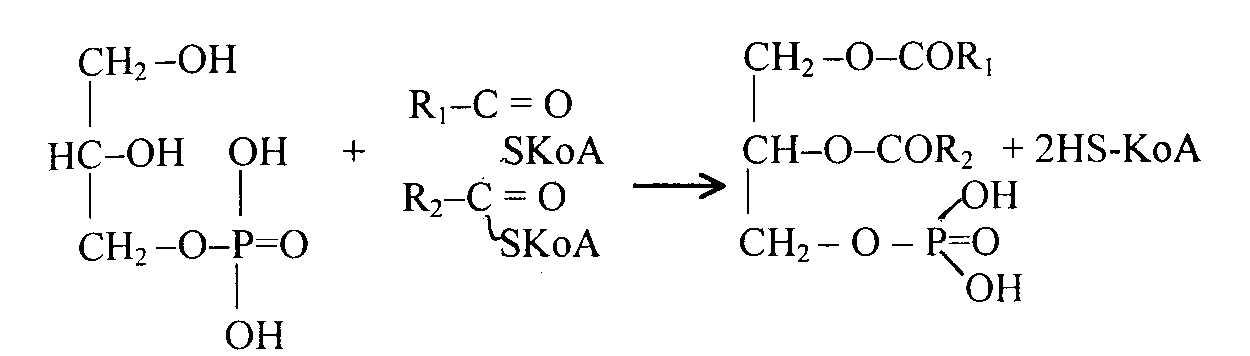
глицерофосфат ацил-КоА фосфатидная кислота
В фосфатидной кислоте в двух позициях спиртовые группы глицерина соединены сложноэфирной связью с остатками жирной кислоты (чаще всего с 16 и 18 углеродными атомами), а в одной позиции с фосфорной кислотой. Синтез триацилглицеринов осуществляется из фосфатидной кислоты. Вначале из фосфатидной кислоты отделяется фосфорная кислота, при этом образуется диацилглицерин. Далее диацилглицерин реагирует с третьей молекулой активированной жирной кислоты (ацил-КоА) и образуется триацилглицерин:
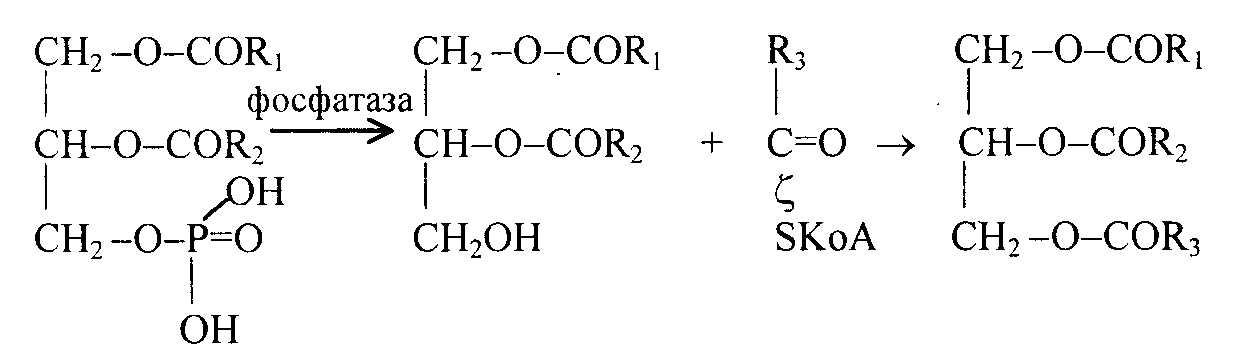
фосфатидная диацилглицерин ацил-КоА триацилглицерин
кислота
Биосинтез фосфоглицеридов происходит в эндо- плазматических мембранах клетки. Фосфатидилэтаноламин синтезируется в следующей последовательности: в начале этаноламин фосфорилируется с участием АТФ и фермента этаноламинкиназы:
![]()
этаноламин фосфоэтаноламин
Затем фосфоэтаноламин реагирует с ЦТФ, в результате образуется цитидилдифосфатэтаноламин и пирофосфат:
фосфоэтаноламин + ЦТФ → ЦДФ-этаноламин.
Далее ЦДФ-этаноламин взаимодействуя с 1,2- диацилглице-ридом, превращается в фосфатидилэтаноламин:
ЦДФ-этаноламин + 1,2-диацилглицерид →
→ фосфатидилэтаноламин + ЦМФ.
Биосинтез фосфатидилхолина (лецитина) осуществляется следующим образом: синтезируется фосфатидилхолин из фосфатидилэтаноламина путем метилирования или же вначале холин взаимодействует с ЦТФ.
Затем ЦДФ-холин с 1,2- диацилглицерином образует дифосфатидилхолин:.
ЦДФ-холин + 1,2 - диацилглицерин →
→ фосфатидилхолин + ЦМФ
Биосинтез фосфатидилсерина происходит путем обмена этаноламина на серин:
Фосфатидилэтаноламин + L-серин →
→ фосфатидилсерин + этаноламин.