

6. Теория климакса, критика ее и современные взгляды на устойчивость и динамику биогеоценозов
Стабильное сообщество, завершающее серию сукцессий (смен), американским экологом Ф. Клементсом (Clements, 1916) названо климаксным, или просто климаксом, находящимся в равновесии с физическим местообитанием. Основные положения теории климакса Ф. Клементса следующие. 1. Основной единицей растительности является формация, которая представляет собой «сложный комплексный организм» («квазиорганизм», по А Тенсли). Подобно организму, формация возникает, растет, зреет и умирает, проходя при этом стадии: детство, юность, зрелое состояние, старость, дряхлость и смерть. 2. Развитие формации протекает в процессе сукцессии от пионерной стадии через промежуточные к конечной, заключительной—к климаксу. 3. Сукцессия с необходимостью идет в направлении от ксерофитной и гигрофитной растительности к мезофитной. Климакс и представляет собой наиболее мезофитный тип, который данный климат может нормально поддерживать. 4. Климаксы изменяются вследствие существенного изменения климата той или иной физико-географической зоны. Такие зональные климаксные сообщества названы климатическими климаксами, а развиваемая Клементсом теория климакса — теорией моноклимакса.
Таким образом, климакс в представлении Ф. Клементса и некоторых его современников за рубежом—это абсолютно устойчивая конечная формация сукцессионной серии, с достижением которой развитие растительности прекращается. Толчком к дальнейшему развитию растительности может стать лишь глубокое изменение климата.
Позднее было установлено, что в пределах той или иной физико-географической зоны наряду с климатическим климаксом наличествует целый ряд климаксных сообществ, связанных с различными формами рельефа, особенностями почвенного покрова и т. п., варьирующих в соответствии с градиентами условий среды. Такого рода климаксные сообщества стали называться субклимаксами. На основе этого сформировалось представление о мозаике климаксных сообществ в пределах зоны или более или менее крупного территориально обособленного географического ландшафта. Это, в свою очередь, послужило основанием для появления теории поликлимакса, нашедшей среди зарубежных биологов немало сторонников.
По Ю. Одуму (1975), стратегия развития экосистемы (биогеоценоза) определяется следующими тремя параметрами: 1) это есть упорядоченный процесс развития сообщества (биоценоза), связанный с изменениями во времени видового состава и протекающих в сообществе процессов, он определенным образом направлен и предсказуем; 2) развитие (или сукцессия) происходит в результате изменений физической среды под действием сообщества; иначе говоря, сукцессия контролируется сообществом, т. е. совокупностью живых компонентов, несмотря на то, что физическая среда определяет характер сукцессии, скорость изменения, а часто и пределы, до которых может дойти развитие; 3) кульминацией развития является стабилизированная экосистема, в которой «на единицу имеющегося потока энергии приходится максимальная биомасса (или высокое содержание информации) и максимальное количество симбиотических связей между организмами» (с. 324).
Заслуживает внимания и следующее замечание Ю. Одума: «Замещение видов в сукцессиях вызывается тем, что популяции, стремясь модифицировать окружающую среду, создают условия, благоприятные для других популяций; это продолжается до тех пор, пока не будет достигнуто равновесие (относительное соответствие.— Л. Н.) между биотическими и абиотическими компонентами» (с. 324). «Одним словом,—заключает автор,—«стратегия» сукцессии, как быстро протекающего процесса, в своей основе сходна со «стратегией» длительного эволюционного развития биосферы: усиление контроля над физической средой (или гомеостаз со средой) в том смысле, что система достигает максимальной защищенности от резких изменений среды» (с. 324).
Для климаксного сообщества, по Ю. Одуму, характерно то, что годовая продукция и приток вещества извне уравновешиваются годовым потреблением, расходом и выносом веществ в окружающее пространство; поэтому чистая годовая продукция в нем отсутствует.
Р. Дажо (1975) приводит ряд закономерностей изменения в ходе сукцессии некоторых свойств биоценозов и экосистем (биогеоценозов) в целом в направлении к климаксному ценозу независимо от типа сукцессии: а) первоначально линейные пищевые цепи с преобладанием растительноядных животных превращаются в сложные пищевые сети, где все большую роль начинают играть детритоядные формы; б) экологические ниши все более сужаются и подвергаются специализации; размеры организмов растут, биологические круговороты удлиняются и усложняются; в) общее количество органических веществ, скопившихся в экосистеме, на первых порах невелико, но неуклонно возрастает, одновременно увеличивается видовое и биохимическое разнообразие; г) отношение «валовая продуктивность—дыхание», обычно превышающее единицу в биоценозах-пионерах, стремится к единице в климаксных биоценозах; д) отношение «валовая продуктивность—биомасса», вначале высокое, начинает уменьшаться, оно соответствует скорости обновления биоценоза; е) отношение «биомасса — энергетический поток», напротив, по мере приближения к климаксному биоценозу увеличивается.
Таким образом, когда выяснилось, что биоценозы и образуемые ими биокосные системы представляют собой открытые системы, непрерывно изменяющиеся во времени, концепция климаксного сообщества, как некоего неизменного «квазиорганизма», подверглась критике и претерпела значительные изменения, а некоторые зарубежные экологи, по утверждению Р. Риклефса, вовсе отказались от нее.
Е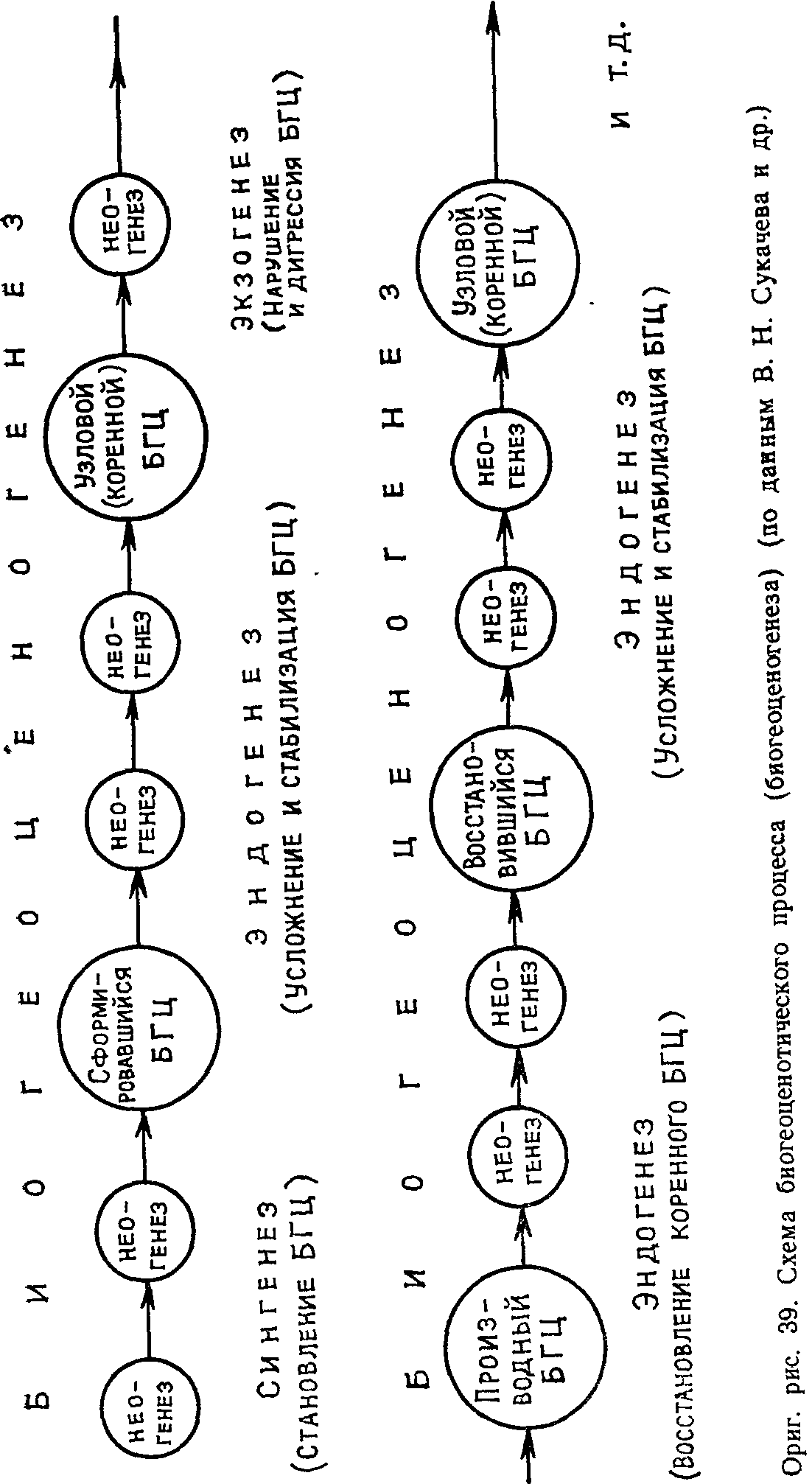 ще
более резкой критике концепция
климакса Ф. Клементса подверглась в
недалеком прошлом со стороны советских
фитоценологов, в особенности
представление о завершающем серию смен
ценозе как заключительном, конечном,
застывшем в неподвижности, дальнейшее
развитие которого возможно лишь под
действием резких климатических перемен
или каких-либо других широкомасштабных
и кардинальных причин (например, внедрение
в ценоз новых, более конкурентомощных
видов и т. п.). Критиковались и такие
стороны ее, как аналогизация развития
ценоза с онтогенезом отдельного организма
и пр. В силу этого советские ценологи
В. Н. Сукачев (1942), П. Д. Ярошенко (1949), А.
П. Шенников (1964) и др. сочли целесообразным
отказаться от термина «климакс»,
заменив его такими терминами, как
выработавшийся или узловой ценоз. Этим
самым подчеркивалось отрицательное
отношение к заложенному в термине
«климакс» представлению об окончании
развития растительного или
биогеоценотического покрова по достижении
этого этапа; с другой стороны, указывалось
на относительный характер стабильности,
устойчивости ценозов на этом этапе.
ще
более резкой критике концепция
климакса Ф. Клементса подверглась в
недалеком прошлом со стороны советских
фитоценологов, в особенности
представление о завершающем серию смен
ценозе как заключительном, конечном,
застывшем в неподвижности, дальнейшее
развитие которого возможно лишь под
действием резких климатических перемен
или каких-либо других широкомасштабных
и кардинальных причин (например, внедрение
в ценоз новых, более конкурентомощных
видов и т. п.). Критиковались и такие
стороны ее, как аналогизация развития
ценоза с онтогенезом отдельного организма
и пр. В силу этого советские ценологи
В. Н. Сукачев (1942), П. Д. Ярошенко (1949), А.
П. Шенников (1964) и др. сочли целесообразным
отказаться от термина «климакс»,
заменив его такими терминами, как
выработавшийся или узловой ценоз. Этим
самым подчеркивалось отрицательное
отношение к заложенному в термине
«климакс» представлению об окончании
развития растительного или
биогеоценотического покрова по достижении
этого этапа; с другой стороны, указывалось
на относительный характер стабильности,
устойчивости ценозов на этом этапе.
В представлении советских ценологов развитие растительного или биогеоценотического покрова с завершением той или иной серии смен фито- или биогеоценозов не прекращается, а замедляется, точнее, протекает как бы в скрытой форме. Растительное сообщество или биогеоценоз в ходе фито- или биогеоценогенеза достигает на стадии «узлового» («выработавшегося») ценоза своего совершенства в компонентном составе, морфологической структуре, функциональной организации, в деятельности механизмов авторегуляции, обеспечивающей слаженное, устойчивое функционирование фитоценоза или биогеоценоза в течение неопределенно длительного времени, поскольку продолжительность их существования не запрограммирована. На этом этапе в фито- или биогеоценозе происходят разнообразные изменения количественного порядка: а) в популяционно-видовом составе живых компонентов за счет внедрения из окружения новых видов и исчезновения старых, оказавшихся менее приспособленными к изменившимся условиям биотопа; б) в структуре и функциональной организации ценозов в связи с изменением количественного соотношения ценозообразующих видов и характера взаимоотношений их между собой в составе каждого ценокомплекса; в) во взаимосвязи биоценоза как совокупности живых компонентов с биоценосредой и всего биогеоценоза с биотическим и абиотическим окружением и пр.
Все эти малозаметные сами по себе
количественные изменения в рамках
существующего «узлового» растительного
сообщества или биогеоценоза, постепенно
накапливаясь, все более и более
нарушают его относительное равновесие,
и далее с течением времени, как отмечает
П. Д. Ярошенко, они становятся настолько
значительными, что выработанность
ценоза, слаженность всех его элементов
совершенно расшатывается. Все э
Рис. 40. Смены ассоциаций пырейника
через ценогенетические ряды эпипарцелл
(по Л. И. Номоконову, 1973). Эпипарцеллы
основные: 1—дербенниково-пырейная;
2 — бекманниево-дербенниково-пырейная.
Эпипарцеллы дополняющие: 3 —
дербенниково-ясколково-пырейная; 4 —
осоково-вербейниково-пырейная;
5—солодково-дербенниково-пырейная; 6
— солодково-мятликово-пырейная; 7 —
бекманниево-жерухово-пырейная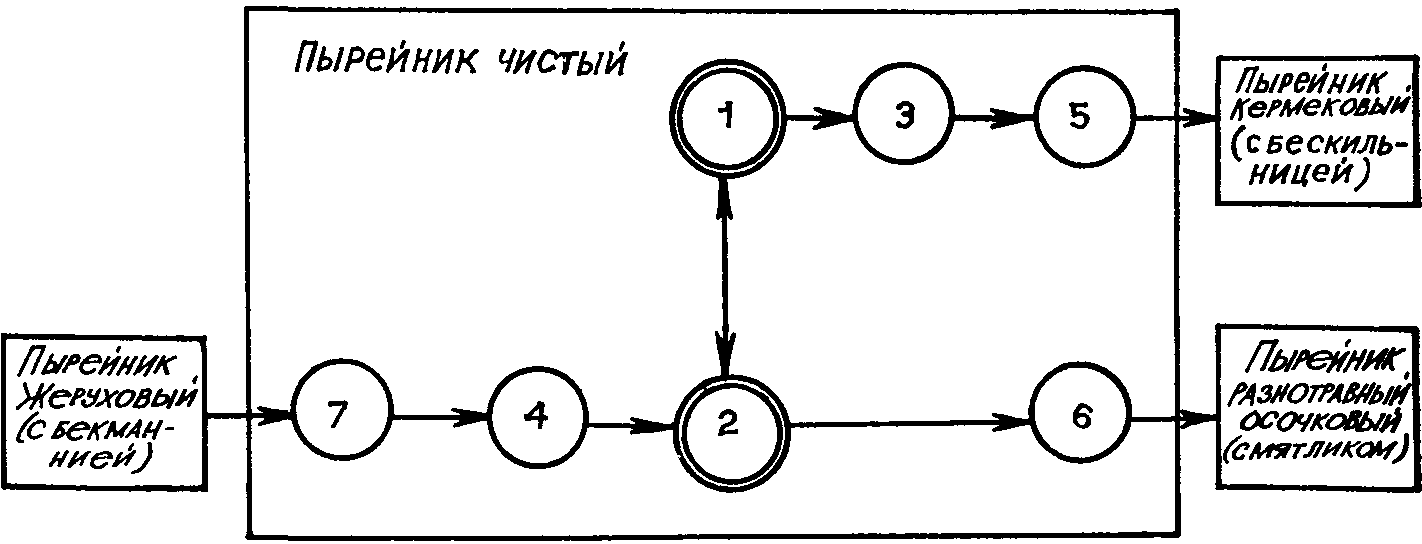 то
в конечном итоге ведет к качественному
изменению «узлового» фито- или
биогеоценоза, т. е. к коренной смене его
иным, новым ценозом, начинающим собой
новую серию смен растительных
сообществ, или биогеоценозов. Следовательно,
«узловой» фито- или биогеоценоз — это
не что иное, как достигшее наибольшего
совершенства и с
то
в конечном итоге ведет к качественному
изменению «узлового» фито- или
биогеоценоза, т. е. к коренной смене его
иным, новым ценозом, начинающим собой
новую серию смен растительных
сообществ, или биогеоценозов. Следовательно,
«узловой» фито- или биогеоценоз — это
не что иное, как достигшее наибольшего
совершенства и с
Ход биогеоценотического процесса (биогеоценогенеза) можно иллюстрировать следующей схемой (рис. 39). На схеме видно, что биогеоценогенез, как общий и относительно медленный и длительный процесс, слагается из более быстрых частных процессов—неогенезов (по В. Б. Сочаве, 1944). Таким образом, биогеоценогенез можно рассматривать как содержание процесса формирования биогеоценоза и последующего его совершенствования и стабилизации; сингенез, эндогенез и экзогенез — как формы проявления этого процесса во времени, а неогенезы — как скоротечные этапы, звенья его. В качестве таких звеньев биогеоценогенеза, как показали наши исследования природных луговых биогеоценозов в пойме Нижнего Дона (Л. И. Номоконов и др., 1978), могут выступать так называемые эпипарцеллы, приуроченные к определенным элементам микрорельефа со свойственными им режимами увлажнения, минерального питания и т. п. Их, следовательно, можно считать временными категориями, развернутыми в пространстве в виде горизонтальных структур биогеоценозов (рис. 40).