

8. Системообразующие единицы, их место и роль в становлении и функциональной организации биогеоценоза
Перейдем к самому главному: к рассмотрению вопроса о формировании биогеоценоза и его функциональной организации из соответствующих составных элементов и функциональных структурных единиц.
«Элементарные функциональные единицы биогеоценоза — это видовые популяции, интегрированное единство которых и составляет биогеоценоз»,—пишет С. С. Шварц (1971, с. 6). Но этот процесс интеграции ценопопуляций в биогеоценоз протекает, судя по всему, через ряд ступеней, представляющих собой качественно различающиеся биологические и ценотические системы разного ранга и соответствующие им формы системного единства, в которые они облекаются. Взяв за основу схему двух параллельных рядов надорганизменной системы иерархии живого (автотрофно-фитоценотического и консортивно-биоценотического), предложенную К. А. Куркиным и А. Р. Матвеевым (1981), мы нашли возможным построить схему, в некотором отношении сходную с ней, но и отличающуюся от нее (табл. 1).
На схеме показаны два взаимосвязанных ряда биологических и ценотических образований разного ранга. Первый ряд состоит из последовательно усложняющихся биологических и ценотических систем: а) особь (организм); б) видовая ценопопуляция; в) ценокомплексы авто-, био- и сапротрофов; г) биота как система живых компонентов. Во втором ряду схемы в такой же последовательности расположены соответствующие каждой био- и ценосистеме формы системного единства: а) индивидуальная консорция; б) популяционная консорция; в) биоценоз как совокупность популяционных консорций; г) биогеоценоз. Нетрудно в каждой ступени этого двоякого рода биологических и ценотических образований увидеть единство двух взаимосвязанных сторон такой парной категории, как содержание и форм а.
И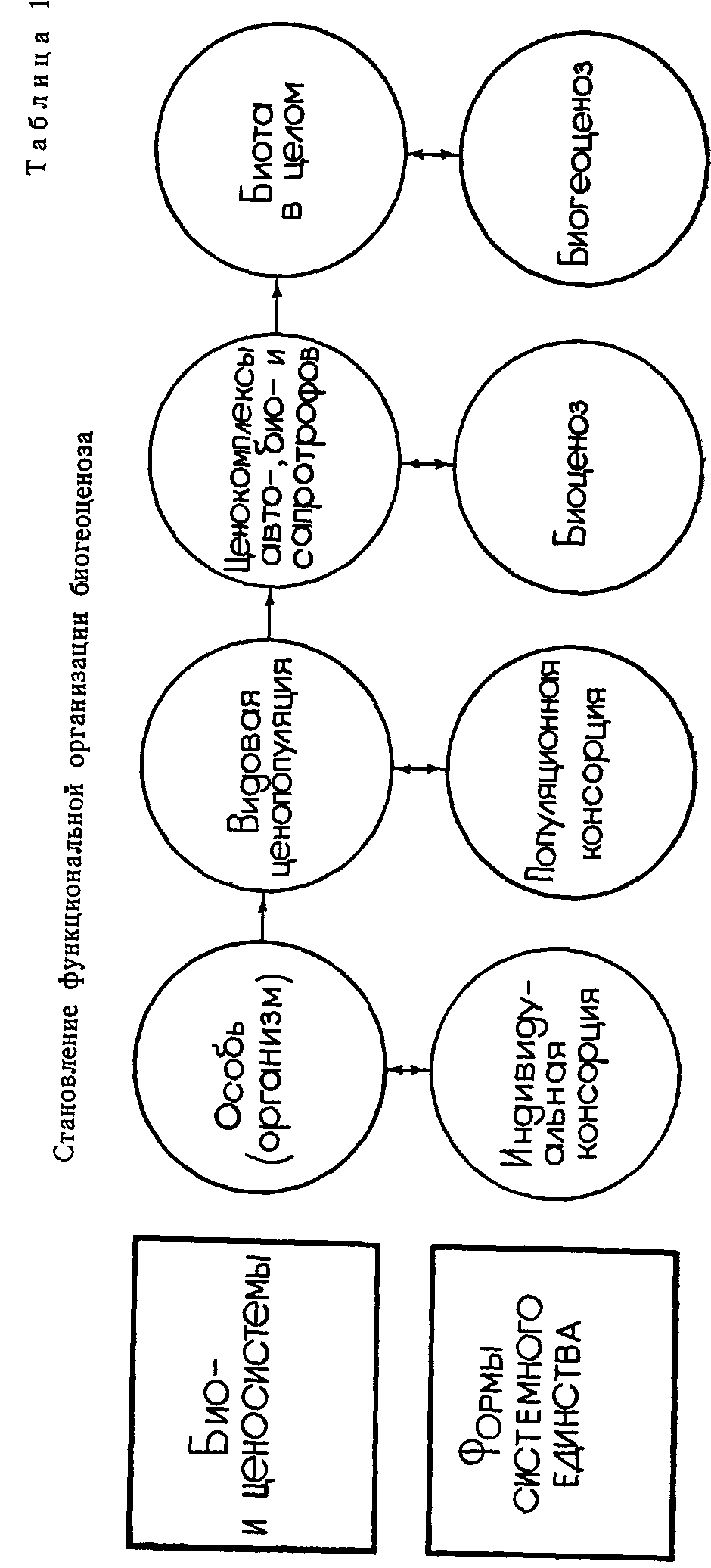 сходя
из такой трактовки существа множества
элементов разного типа и ранга,
рассматриваемых многими в качестве
функциональных структурных единиц
функциональной организации биогеоценоза,
представляется возможным, на наш взгляд,
с большей точностью определить
действительное место и ранг каждого из
них в общей иерархии системообразующих
единиц, а также соотношение существующих
взаимосвязей их между собой. Так,
доминирующие особи автотрофов, являясь
центрами индивидуальных консорций,
входят в качестве элементов в виде
экобиоморф (иначе— фитоценотипов) в
состав видовых ценопопуляций автотрофов,
участвуя при этом вместе со своей
«свитой» разнообразных консортов в
формировании соответствующих популяционных
консорций— полночленных (пастбищных,
биотрофных) и неполночленных (детритных,
сапротрофных). Ценозообразующие видовые
ценопопуляции, являющиеся системами
особей живых существ и одновременно
центрами или консортами популяционных
консорций,, входят в качестве элементов
в состав автотрофных, биотрофных и
сапротрофных ценокомплексов, занимая
в каждом из них определенные позиции в
трофических звеньях пастбищной или
детритной пищевой цепи. Ценокомплексы
авто-, био- и сапротрофов,, являющихся
системами соответствующих ценозообразующих
видовых ценопопуляций и одновременно
живыми компонентами биоценоза, входят
в качестве элементов в состав биоты,
выполняя присущие им функции в обмене
веществ и преобразовании энергии в
трофических звеньях пищевых цепей и
сетей. Биота, являющаяся целостной
системой авто-, био- и сапротрофного
ценокомплексов и одновременно подсистемой
биогеоценоза, входит в форме биоценоза
в его состав; второй подсистемой
биогеоценоза как основной
структурно-функциональной единицы
биогеосферы является биотоп.
сходя
из такой трактовки существа множества
элементов разного типа и ранга,
рассматриваемых многими в качестве
функциональных структурных единиц
функциональной организации биогеоценоза,
представляется возможным, на наш взгляд,
с большей точностью определить
действительное место и ранг каждого из
них в общей иерархии системообразующих
единиц, а также соотношение существующих
взаимосвязей их между собой. Так,
доминирующие особи автотрофов, являясь
центрами индивидуальных консорций,
входят в качестве элементов в виде
экобиоморф (иначе— фитоценотипов) в
состав видовых ценопопуляций автотрофов,
участвуя при этом вместе со своей
«свитой» разнообразных консортов в
формировании соответствующих популяционных
консорций— полночленных (пастбищных,
биотрофных) и неполночленных (детритных,
сапротрофных). Ценозообразующие видовые
ценопопуляции, являющиеся системами
особей живых существ и одновременно
центрами или консортами популяционных
консорций,, входят в качестве элементов
в состав автотрофных, биотрофных и
сапротрофных ценокомплексов, занимая
в каждом из них определенные позиции в
трофических звеньях пастбищной или
детритной пищевой цепи. Ценокомплексы
авто-, био- и сапротрофов,, являющихся
системами соответствующих ценозообразующих
видовых ценопопуляций и одновременно
живыми компонентами биоценоза, входят
в качестве элементов в состав биоты,
выполняя присущие им функции в обмене
веществ и преобразовании энергии в
трофических звеньях пищевых цепей и
сетей. Биота, являющаяся целостной
системой авто-, био- и сапротрофного
ценокомплексов и одновременно подсистемой
биогеоценоза, входит в форме биоценоза
в его состав; второй подсистемой
биогеоценоза как основной
структурно-функциональной единицы
биогеосферы является биотоп.
Итак, все иерархически последовательно взаимосвязанные биологические и ценотические составные элементы функциональной организации биогеоценоза и соответствующие им формы системного единства оказываются задействованными. При этом каждый из них занимает определенное место в общей системе биогеоценоза, обеспечивающее выполнение свойственной ему функции.
Из изложенного следует, что формирование биогеоценоза и его функциональной организации—многоступенчатый процесс, который совершается при непосредственном участии различных по типу питания и использованию источников энергии видовых ценопопуляций, объединенных в соответствующие ценокомплексы, в совокупности образующие биоту. Популяционные же консорций, как и пищевые цепи (и сети) в их составе, с одной стороны, биоценозы и биогеоценозы—с другой,—это не что иное, как формы, выражающие весьма сложные, многократно переплетающиеся связи и отношения видовых ценопопуляций автотрофного, биотрофного и сапротрофного ценокомплексов и биоты в целом. Занимая в их составе определенные позиции, они включаются, таким образом, в форме соответствующих трофических уровней в состав пищевых цепей той или иной популяционной консорций. Последняя же с позиции системной трактовки организации биогеоценоза представляет собою функциональную структурную единицу более крупного системного единства-биоценоза, в форме которого биота входит в качестве подсистемы в состав биогеоценоза как материально-энергетической ячейки биогеосферы.
Таким образом, несколько перефразируя отнесенные к фитоценозу слова А. П. Шенникова (1964), можно сказать, что биоценоз и биогеоценоз в целом есть оформление бесконечного многообразия взаимных связей и отношений их составных элементов разного ранга.
Так в общей форме представляется становление биогеоценоза и формирование его функциональной организации из соподчиненных биологических и ценотических образований разного ранга и соответствующих форм системного единства. Более конкретно этот процесс представлен П. М. Рафесом (1972) на материале исследования осиново-березовых рощиц в «блюдцах» лесостепной полосы Западной Сибири, называемых «березовыми колками». Рассматривая биогеоценоз как систему консорций, автор обращает внимание на то обстоятельство, что в биогеоценозе, как правило, действуют несколько популяционных консорций, детерминированных определенными ценопопуляциями автотрофных растительных доминант, например березой, осиной, сосной и др. Вследствие количественного и качественного различия продуцируемой названными детерминантами фитомассы подбирается и различный состав консортов I концентра в каждой из них, что определяет и состав консортов последующих концентров консорций. Из этого им делается вывод: биогеоценоз нельзя изображать как систему, характеризующуюся единым потоком веществ и энергии. В действительности он складывается из нескольких вещественно-энергетических потоков, входящих в состав биогеоценоза консорций. Более того, поскольку консорций включают по несколько пищевых цепей, каждая из которых также характеризуется специфичным потоком веществ и заключенной в них энергии, то количество и разнообразие вещественно-энергетических потоков еще более возрастает. «В конечном счете,—заключает автор,—функция биогеоценоза — это осуществление круговорота материи на занимаемой им территории» (с. 45).
И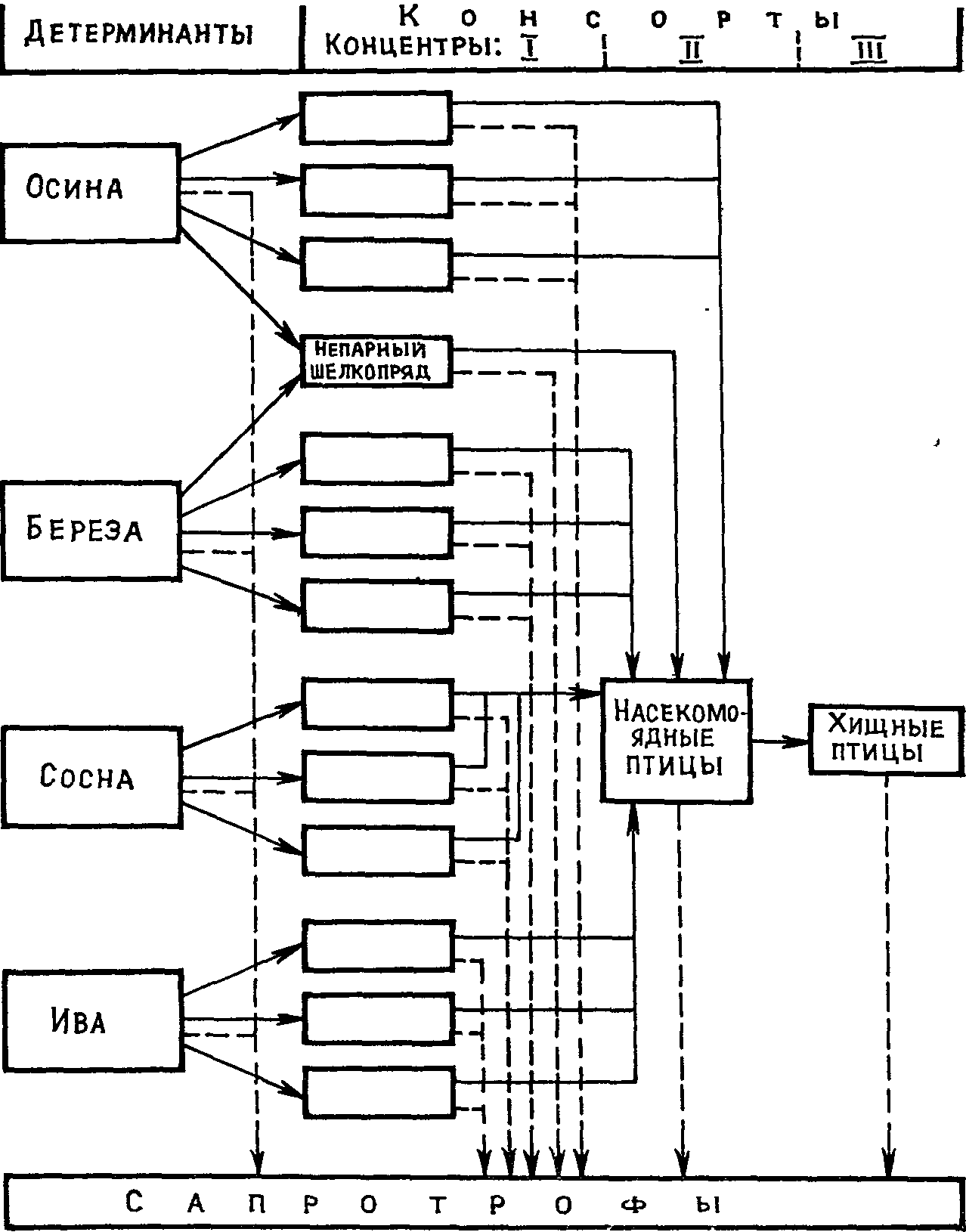 зложенное
представление автора о формировании
биогеоценоза и его функциональной
организации из слагающих его популяционных
консорций весьма наглядно иллюстрируется
схемой (рис. 19). Заслуживает внимания и
оригинальная схема консорций березы
(рис. 20), составленная автором не в виде
концентрических кругов вокруг
детерминанта, а в виде развернутой в
одном направлении полосы. Это дало
возможность более полно показать большое
разнообразие консортивных связей между
консортами и консортов с центром
консорций, в том числе и связь потребителей
живой биомассы всех концентров и
детерминанта консорций с сапротрофами,
т. е. показано завершение цикла обмена
веществ в рамках популяционной консорций.
зложенное
представление автора о формировании
биогеоценоза и его функциональной
организации из слагающих его популяционных
консорций весьма наглядно иллюстрируется
схемой (рис. 19). Заслуживает внимания и
оригинальная схема консорций березы
(рис. 20), составленная автором не в виде
концентрических кругов вокруг
детерминанта, а в виде развернутой в
одном направлении полосы. Это дало
возможность более полно показать большое
разнообразие консортивных связей между
консортами и консортов с центром
консорций, в том числе и связь потребителей
живой биомассы всех концентров и
детерминанта консорций с сапротрофами,
т. е. показано завершение цикла обмена
веществ в рамках популяционной консорций.
В
Рис. 19. Схема функциональной
организации биогеоценоза (по П. М.
Рафесу, 1972)