

57 Вопрос
Глюконеогенез, ключевые ферменты, значение в метаболизме плода. Регуляция гликолиза и глюконеогенеза в печени. Цикл Кори. Глюкозо-аланиновый цикл.
Глюконеогенез – путь образования глюкозы denovo из низкомолекулярных метаболитов углеводного обмена цикла Кребса и соединений неуглеводной природы.( молочная и пи-ровиноградная кислоты, так называемые гликогенныеаминокислоты, глицерол) Поддерживает уровень глюкозы в крови в период длительного голодания и интесивных физических нагрузок.Локализуется в печени,в корковом веществе почек, в слизистый оболочке кишечника.
Образование фосфоенолпирувата из пирувата. Синтез фосфоенолпирувата осуществляется в несколько этапов. Первоначально пируват под влиянием пируваткарбоксилазыи при участии СО2 иАТФкарбоксилируется с образованием оксалоацетата:

Затем оксалоацетат в результате декарбоксилированияи фосфорилирования под влияниемферментафосфоенолпируваткарбоксилазы превращается в фосфоенолпируват.Доноромфосфатного остатка вреакциислужит гуанозинтрифосфат (ГТФ):
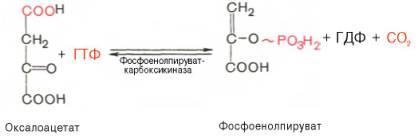
Установлено, что в процессе образования фосфоенолпирувата участвуют ферментыцитозоля имитохондрий.
Первый этап синтеза протекает в митохондриях(рис. 10.6). Пируват-карбоксилаза, которая катализирует этуреакцию, является аллостери-ческим митохондриальнымферментом. В качестве аллостерическогоактиватораданногоферментанеобходим ацетил-КоА.Мембранамитохондрийнепроницаема для образовавшегося оксалоацетата. Последний здесь же, вмитохондриях, восстанавливается в малат:

Реакцияпротекает при участии митохондриальной НАД-зависимоймалатдегидрогеназы. Вмитохондрияхотношение НАДН/НАД+ относительно велико, в связи с чем внутримитохондриальный оксалоацетат легко восстанавливается в малат, который легко выходит измитохондриичерез митохондриальнуюмембрану. В цитозоле отношение НАДН/НАД+ очень мало, и малат вновь окисляется при участии цитоплазматической НАД-зависимоймалатдегидрогеназы:
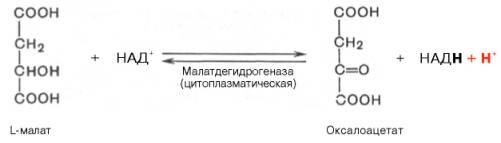
Дальнейшее превращение оксалоацетата в фосфоенолпируват происходит в цитозоле клетки.
Превращение фруктозо-1,6-бисфосфата во фруктозо-6-фосфат. Фосфо-енолпируват, образовавшийся из пирувата, в результате ряда обратимых реакцийгликолизапревращается во фруктозо-1,6-бисфосфат. Далее следует фосфофруктокиназнаяреакция, которая необратима.Глюконеогенезидет в обход этой эндергоническойреакции. Превращение фруктозо-1,6-бис-фосфата во фруктозо-6-фосфат катализируется специфическойфосфатазой:
![]()
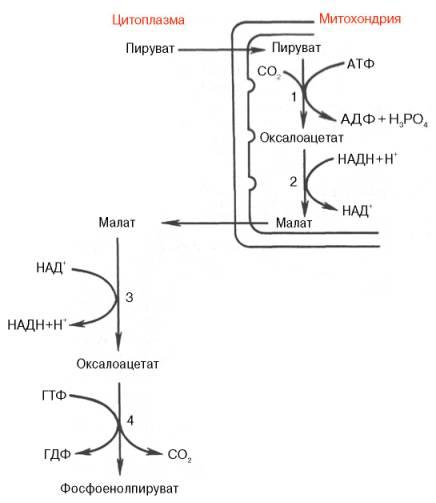
Рис. 10.6. Образование фосфоенол-пирувата из пирувата. 1 - пируваткарбоксилаза; 2 - малатде-гидрогеназа (митохондриальная); 3 -малатдегидрогеназа (цитоплазматиче-ская); 4 - фосфоенолпируват-карбокси-киназа.
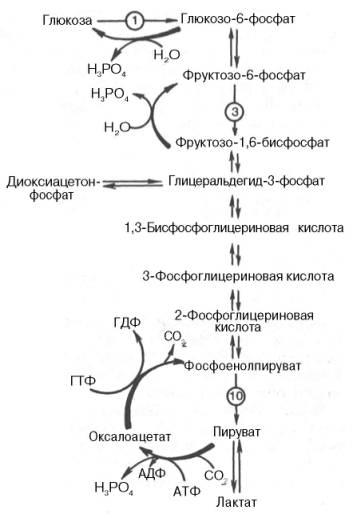
Образование глюкозыиз глюкозо-6-фосфата. В последующей обратимой стадиибиосинтезаглюкозыфруктозо-6-фосфат превращается в глюкозо-6-фосфат. Последний может дефосфорилироваться (т.е.реакцияидет в обход гексокиназнойреакции) под влияниемферментаглюкозо-6-фосфатазы:
![]()
Регуляция глюконеогенеза. Важным моментом в регуляции глюконеогенеза является реакция, катализируемаяпируваткарбоксилазой. Роль положительного аллостерического модулятора этогоферментавыполняет ацетил-КоА. В отсутствие ацетил-КоАферментпочти полностью лишенактивности. Когда вклеткенакапливается митохондриальный ацетил-КоА,биосинтезглюкозыиз пирувата усиливается. Известно, что ацетил-КоА одновременно является отрицательным модулятором пируватдегидрогеназного комплекса. Следовательно, накопление ацетил-КоА замедляет окислительноедекарбоксилированиепирувата, что также способствует превращению последнего вглюкозу.
Другой важный момент в регуляции глюконеогенеза–реакция, катализируемая фруктозо-1,6-бисфосфатазой –ферментом, который ингибируетсяАМФ. Противоположное действиеАМФоказывает на фосфофруктокиназу, т. е. для этогоферментаон является аллостерическимактиватором. При низкойконцентрацииАМФи высоком уровнеАТФпроисходит стимуляцияглюконеогенеза. Напротив, когда величина отношенияАТФ/АМФмала, вклеткенаблюдается расщеплениеглюкозы.
В 1980 г. группой бельгийских исследователей (Г. Херс и др.) в тканипеченибыл открыт фруктозо-2,6-бисфосфат, который является мощным регуляторомактивностидвух перечисленныхферментов:
Фруктозо-2,6-бисфосфат
активирует фосфофруктокиназу и ингибирует
фруктозо-1,6-бисфосфатазу. Повышение
вклеткеуровня
фруктозо-2,6-бис-фосфата способствует
усилениюгликолизаи
уменьшению скорости глю-конеогенеза.
При сниженииконцентрациифруктозо-2,6-бисфосфата
отмечается обратная картина.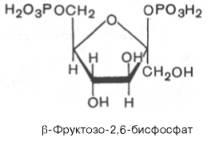
Установлено, что биосинтезфруктозо-2,6-бисфосфата происходит из фруктозо-6-фосфата при участииАТФ, а распадается он на фруктозо-6-фосфат инеорганический фосфат.Биосинтези распад фруктозо-2,6-бис-фосфата катализируется одним и тем жеферментом, т.е. данныйферментбифункционален, он обладает и фосфокиназной, и фосфатазнойактивностью:
Показано
также, что бифункциональный ферментв
свою очередь регулируется путем
цАМФ-зависимогофосфорилирования.
Фосфорилирова-ние приводит к увеличению
фосфатазнойактивностии
снижению
фосфо-киназнойактивностибифункциональногофермента.
Этот механизм объясняет быстрое
воздействиегормонов,
в частностиглюкагона,
на уровень фруктозо-2,6-бисфосфата
вклетке.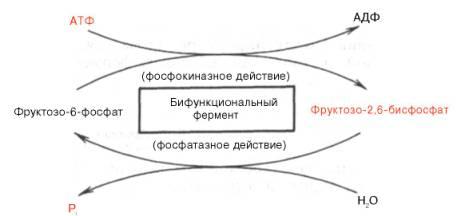
Активностьбифункциональногоферментарегулируется также некоторымиметаболитами, среди которых наибольшее значение имеет гли-церол-3-фосфат. Действие глицерол-3-фосфата наферментпо своей направленности аналогично эффекту, который наблюдается при его фосфорилировании с помощью цАМФ-зависимыхпротеинкиназ.
В настоящее время фруктозо-2,6-бисфосфат, помимо печени, обнаружен и в других органах итканяхживотных, а также у растений имикроорганизмов.
Показано, что глюконеогенезможет регулироваться и непрямым путем, т.е. через изменениеактивности фермента, непосредственно не участвующего в синтезеглюкозы. Так, установлено, чтоферментгликолизапируваткиназа существует в 2 формах – L и М. Форма L (от англ. liver –печень) преобладает втканях, способных кглюконеогенезу. Эта форма ингибируется избыткомАТФи некоторымиаминокислотами, в частности ала-нином. М-форма (от англ. muscle – мышцы) такой регуляции не подвержена. В условиях достаточного обеспеченияклеткиэнергией происходит ингибирование L-формыпируваткиназы. Как следствие ингибирования замедляетсягликолизи создаются условия, благоприятствующие глюконеогенезу.
Наконец, интересно отметить, что между гликолизом, интенсивно протекающим вмышечной тканипри ее активной деятельности, и глюконеогенезом, особенно характерным для печеночнойткани, существует тесная взаимосвязь. При максимальнойактивностимышц в результате усилениягликолизаобразуется избытокмолочной кислоты, диффундирующей вкровь, впеченизначительная ее часть превращается вглюкозу(глюконеогенез). Такаяглюкозазатем может быть использована как энергетическийсубстрат, необходимый для деятельностимышечной ткани. Взаимосвязь между процессамигликолизавмышечной ткании глюконеогенезом впечениможет быть представлена в виде схемы:
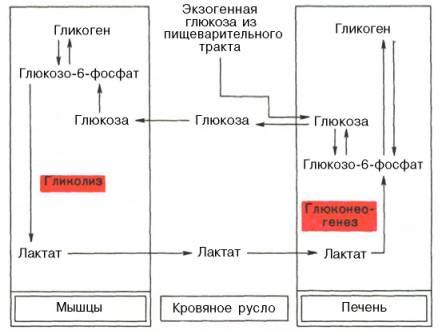
1. 2.
2.
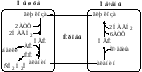
1.Глюкозо-лактатный цикл(цикл Кори) не требует наличие кислорода, функционирует всегда, обеспечивает: 1) утилизацию лактата, образующегося в анаэробных условиях (скелетные мышцы, эритроциты), что предотвращает лактоацидоз; 2) синтез глюкозы (печень).
2.Глюкозо-аланиновый цикл функционирует в мышцах при голодании. При дефиците глюкозы, АТФ синтезируется за счет распад белков и катаболизма аминокислот в аэробных условиях, при этом глюкозо-аланиновый цикл обеспечивает: 1) удаление азота из мышц в нетоксичной форме; 2) синтез глюкозы (печень).
Цикл Кори(глюкозолактатный цикл)
Лактат, образовавшийся в интенсивно работающих мышцах или в клетках с преобладающим анаэробным способом катаболизма глюкозы, поступает в кровь, а затем в печень .в печени отношение NADH/NAD ниже, чем в сокращающейся мышце, поэтому лактатдегидрогеназная реакция протекает в обратном направление, т.е. в сторону образования пирувата из лактата. Далее пируват включается в глюконеогенез, а образовавшаяся глюкоза поступает в кровь и поглащается скелетным мышцами.Цикл Кори включает 2 важные функции:
1-обеспечивает утилизация лактата.
2-предотвращает накопление лактата.
Глюкозо-аланиновый цикл:
Глюкоза в мышцах – пируват в мышцах- аланин в мышцах-аланин в печени – глюкоза в печени – глюкоза в мышцах.
Весь цикл не приводит к увеличению количества глюкозы в мышцах,но он решает проблемы транспорта аминного азота из мышц в печень и предотвращает лактоацидоз.