

45 Вопрос
Пути биосинтеза заменимых аминокислот: реакции, ферменты.
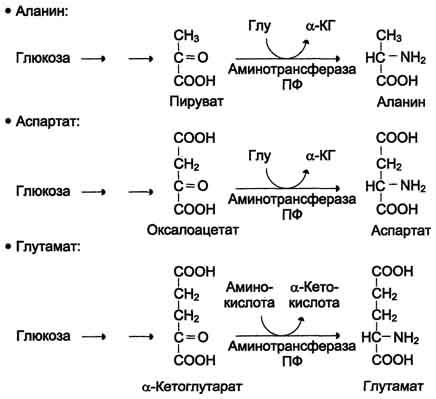
В организме человека возможен синтез восьми заменимых аминокислот: Ала, Асп, Асн, Сер, Гли, Глу, Глн, Про (рис. 9-23). Углеродный скелет этих аминокислот образуется из глюкозы. α-Аминогруппа вводится в соответствующие α-кетокислоты в результате реакций трансаминирования.Универсальным донором α-аминогруппы служит глутамат.
Путём трансаминирования α-кетокислот, образующихся из глюкозы, синтезируются аминокислоты (см. схему А на с. 492).
Глутамат также образуется при восстановительном аминировании α-кетоглутарата глутаматдегидрогеназой.
Эти реакции обратимы и играют большую роль как в процессе синтеза аминокислот, так и при их катаболизме. Такие реакции, выполняющие двойную функцию, называют амфиболическими.
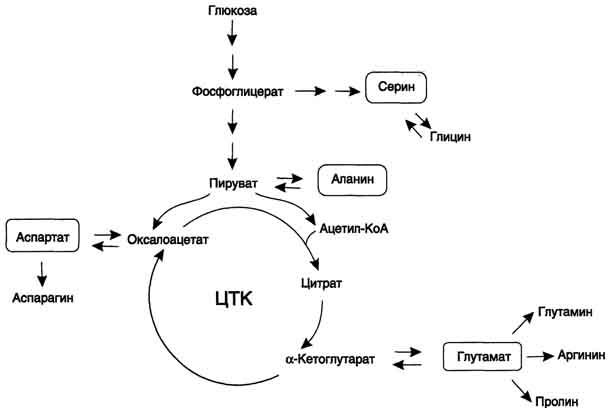
Рис. 9-23. Пути биосинтеза заменимых аминокислот.
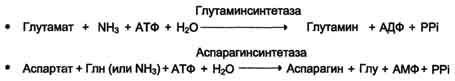
Амиды
глутамин и аспарагин синтезируются
из соответствующих дикарбоновых
аминокислот Глу и Асп (см. схему А).
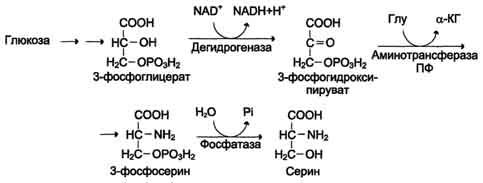
Серин образуется из 3-фосфоглицерата - промежуточного продукта гликолиза, который окисляется до 3-фосфопирувата и затем трансаминируется с образованием се-рина (см. схему Б).
Существует 2 пути синтеза глицина:
1) из серина с участием производного фолиевой кислоты в результате действия се-риноксиметилтрансферазы:
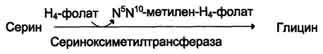
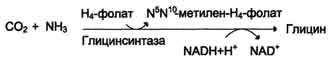
2) в результате действия фермента глицинсинтазы в реакции:
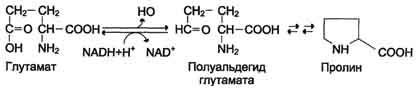
Пролин синтезируется из глутамата в цепи обратимых реакций. Эти же реакции используются и при катаболизме пролита (см. схему В на с. 494).
Кроме восьми перечисленных заменимых аминокислот, в организме человека могут синтезироваться ещё четыре аминокислоты.
Частично заменимые аминокислоты Apr и Гис синтезируются сложным путём в небольших количествах. Большая их часть должна поступать с пищей.
Синтез аргинина происходит в реакциях орнитинового цикла (см. выше подраздел IV);
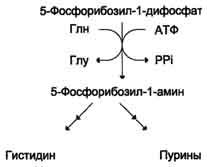
Гистидин синтезируется из АТФ и рибозы. Часть имидазольного цикла гистидина - N=CH-NH- образуется из пуринового ядра аденина, источником которого служит АТФ, остальная часть молекулы - из атомов рибозы. При этом образуется 5-фосфорибозиламин, который кроме синтеза гистидина необходим для синтеза пуринов.
Для синтеза условно заменимых аминокислот тирозина и цистеина требуются незаменимые аминокислоты фенилаланин и метионин соответственно (см. подразделы VIII и IX).
Образование других аминокислот также возможно при наличии соответствующих α-кетокислот, которые могут трансаминироваться с глутаматом. Таким образом, незаменимой частью молекулы аминокислот является их углеродный скелет. Источником таких незаменимых ос-кетокислот служат только белки пищи. Исключение составляют лизин и треонин, которые не подвергаются трансаминированию, их а-кетоаналоги с пищей практически не поступают и в организме не синтезируются. Единственный источник этих аминокислот - пищевые белки.
46 Вопрос
Пути обмена серина и глицина. Роль серина, глицина и метионина в образовании одноуглеродных фрагментов и реакциях трансметилирования, участие тетрагидрофолиевой кислоты в этих процессах. Недостаточность фолиевой кислоты. Механизм бактериостатического действия сульфаниламидных препаратов. Особенности обмена метионина. S-аденозилметионин-донор метильных групп.
Обмен серина и глицина
Серин - заменимая аминокислота, синтезируется из промежуточного продукта гликолиза - 3-фосфоглицерата, а аминогруппу получает от глутаминовой кислоты.
Глицин - также заменимая аминокислота, основным источником которой служит серии. Реакцию синтеза глицина из серина катализирует фермент серин-оксиметилтрансфераза, кофермен-том которой является Н4-фолат (см. схему А).
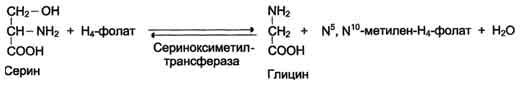
Реакция превращения серина в глицин легко обратима. Основной путь катаболизма глицина у человека и других позвоночных также связан с использованием Н4-фолата (см. схему Б).
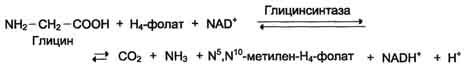
Эта реакция обратима и катализируется глицинсинтазой - ферментным комплексом, похожим на пируватдегидрогеназный комплекс, и локализованным в митохондриях клеток печени. По последним данным глицинрасщепляющая ферментная система несколько отличается от глицинсинтазы и содержит 4 белка: Р-белок, включающий кофермент ПФ, Н-белок, содержащий липоевую кислоту, Т-белок с коферментом Н4-фолат, L-белок, являющийся дигидролипоилдегидрогеназой с коферментом NAD+.