

54 Вопрос
Аэробный гликолиз, последовательность реакций до образования пирувата. Распространение, энергетическая эффективность и физиологическое значение аэробного распада глюкозы. Использование глюкозы для синтеза жиров в печени и жировой ткани.
гликолизпротекает в гиалоплазме (цитозоле)клетки.
1)Начинается с фосфорилирования глюкозы,катализирует гексокиназа
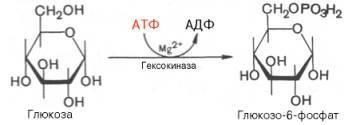
2)изомеризация г-6-ф во ф-6-ф (фермент глюкозо-6-фосфатизомераза.)
Второй реакциейгликолизаявляется превращение глюкозо-6-фос-фата под действиемферментаглюкозо-6-фосфатизомеразы во фруктозо-6-фосфат:

Эта реакцияпротекает легко в обоих направлениях, и для нее не требуется каких-либо кофакторов.
3)фосфорилирование ф-6-ф с образованием фруктозо-1,6-бисфосфата (фермент фосфофруктокиназа. Третья реакциякатализируетсяферментомфосфофруктокиназой; образовавшийся фруктозо-6-фосфат вновь фосфорилируется за счет второймолекулыАТФ:
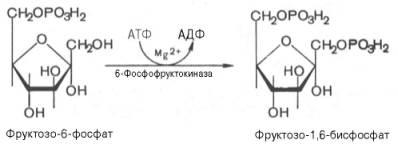
Данная реакцияаналогично гексокиназной практически необратима, протекает в присутствииионовмагнияи является наиболее медленно текущейреакциейгликолиза. Фактически этареакцияопределяет скоростьгликолизав целом.
4)расщепление фруктозо-1,6-бисфосфата на глицеральдегид-3-фосфат и дигидроксиацетат (фермент фруктозо-1,6-бисфосфат-альдолаза). Четвертую реакциюгликолизакатализируетферментальдолаза. Под влиянием этогоферментафруктозо-1,6-бисфосфат расщепляется на две фосфотриозы:
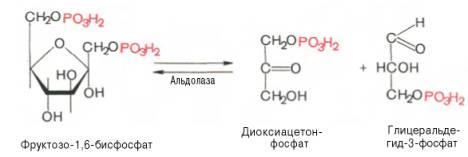
Эта реакцияобратима.5)взаимопревращение триозофосфатов (фермент триозофосфатизомераза) Пятая реакция– этореакцияизомеризациитриозофосфатов. Катализируетсяферментомтриозофосфатизомеразой:

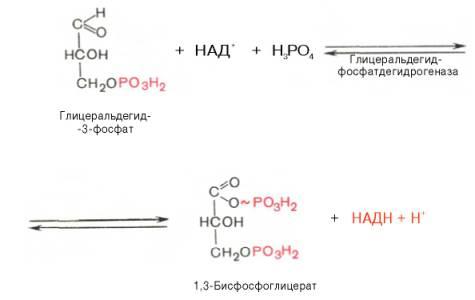
6)окисление глицеральдегид-3-фосфата до 1,3-дифосфоглицерата (фермент глицеральдегид-3-фосфат-дегидрогеназа). В результате шестой реакцииглицеральдегид-3-фосфат в присутствииферментаглицеральдегидфосфатдегидрогеназы,коферментаНАД инеорганического фосфатаподвергается своеобразномуокислениюс образованием 1,3-бисфосфоглицериновойкислотыи восстановленной формы НАД (НАДН). Этареакцияблокируется йод- или бромацетатом, протекает в несколько этапов:
7)перенос фосфатной группы с 1.3-дифосфоглицерата на АДФ (ферм фосфоглицераткиназа) Седьмая реакциякатализируетсяфосфоглицераткиназой, при этом происходит передача богатого энергией фосфатного остатка (фосфатной группы в положении 1) наАДФс образованиемАТФи 3-фосфогли-цериновойкислоты(3-фосфоглицерат):
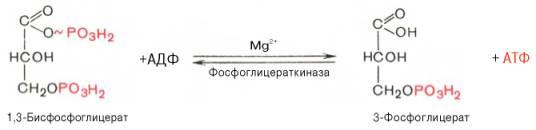
8)изомеризация 3-фосфоглицерата в 2-фосфоглицерат (ферм фосфоглицерат-фосфомутаза) Восьмая реакциясопровождается внутримолекулярным переносом оставшейся фосфатной группы, и 3-фосфоглицериноваякислотапревращается в 2-фосфоглицериновуюкислоту(2-фосфоглицерат).

9)дегидратация 2-фосфоглицерата с обр-м фосфоенолпирувата (ферм енолаза). Девятая реакциякатализируетсяферментоменолазой, при этом 2-фосфоглицериноваякислотав результате отщеплениямолекулыводыпереходит в фосфоенолпировинограднуюкислоту(фосфоенолпируват), а фосфатная связь в положении 2 становится высокоэргической:
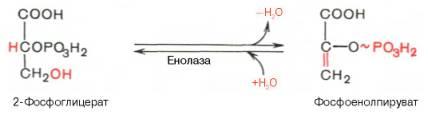
Енолазаактивируется двухвалентнымикатионамиMg2+или Мn2+ и ингибируетсяфторидом.10)перенос фосфатной группы с фосфоенолпирувата на АДФ (ферм пируваткиназа) с образованием пирувата. Десятая реакцияхарактеризуется разрывом высокоэргической связи и переносом фосфатного остатка от фосфоенолпирувата наАДФ(субстратноефосфорилирование). Катализируетсяферментомпируваткиназой:
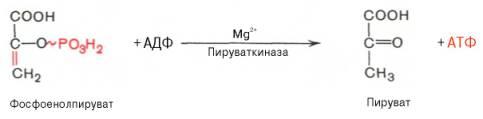
Для действия пируваткиназынеобходимыионыMg2+, а также одновалентныекатионыщелочных металлов(К+ или др.). Внутриклеткиреакцияявляется практически необратимой.
В аэробном гликолизе образуется 10 моль АТР на 1 моль глюкозы. Так, в реакциях 7, 10 образуется 4 моль АТР путем субстратного фосфорилирования, а в реакции 6 синтезируется 6 моль АТР (на 2 моль глицероальдегидфосфата) путем окислительного фосфорилирования. Суммарный эффект аэробного гликолиза составляет 8 моль АТР, так как в реакциях 1 и 3 используется 2 моль АТР. Дальнейшее окисление двух моль пируват в общих путях катаболизма сопровождается синтезом 30 моль АТР ( по 15 моль на каждую молекулу пирувата . Следовательно, суммарный энергетический эффект аэробного распада глюкозы до конечных продуктов составляет 38 моль АТР. Суммарное уравнение С6Н12О6 + 6О2 → 6СО2 + Н2О + 2880кДж/моль. Основное физиологическое значение аэробного распада глюкозы заключается в использовании ее энергии для синтеза АТФ. В наибольшей зависимости от аэробного гликолиза находится мозг. Он расходует 100 г глюкозы в сутки. В состоянии основного обмена около 20% кислорода потребляется мозгом. Поэтому недостаток глюкозы или кислорода проявляется, прежде всего, симптомами со стороны центральной нервной системы - головокружением, потерей сознания, судорогами.
Синтез триглицеридов происходит из глицеринаижирных кислот(главным образом стеариновой, пальмитиновой и олеиновой). Путьбиосинтезатриглицеридов втканяхпротекает через образование α-глице-рофосфата (глицерол-3-фосфата) как промежуточного соединения.
В почках,печени а также в стенке кишечника, где активность ферментаглицеролкиназы(только в печени) высока,глицеринфосфорилируется за счетАТФс образованием глицерол-3-фосфата:
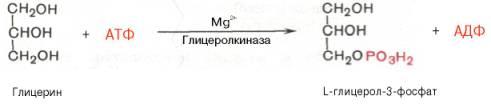
В жировой ткании мышцах вследствие очень низкойактивностиглицеролкиназы образование глицерол-3-фосфата в основном связано с процессамигликолизаигликогенолиза. Известно, что в процессе гли-колитического распадаглюкозыобразуется дигидроксиацетонфосфат (см. главу 10). Последний в присутствии цитоплазматической глицерол-3-фосфатдегидрогеназы способен превращаться в глицерол-3-фосфат:

Отмечено, что если содержание глюкозывжировой тканипонижено (например, при голодании), то образуется лишь незначительное количество глицерол-3-фосфата и освободившиеся в ходе липолиза свободныежирные кислотыне могут быть использованы для ресинтеза триглицеридов, поэтомужирные кислотыпокидаютжировую ткань. Напротив, активациягликолизавжировой тканиспособствует накоплению в ней триглицеридов, а также входящих в их составжирных кислот. Впеченинаблюдаются оба пути образования глицерол-3-фосфата.
Образовавшийся тем или иным путем глицерол-3-фосфат последовательно ацилируется двумя молекуламиКоА-производногожирной кислоты(т.е. «активными» формамижирной кислоты– ацил-КоА). В результате образуется фосфатиднаякислота(фосфатидат):
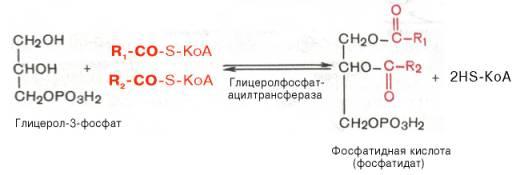
Как отмечалось, ацилированиеглицерол-3-фосфата протекает последовательно, т.е. в 2 этапа. Сначала глицерол-3-фосфат-ацилтрансфераза катализирует образование лизофосфатидата (1-ацилглицерол-3-фосфата, а затем 1-ацилглицерол-3-фосфат-ацилтрансфераза катализирует образование фосфатидата (1,2-диацилглицерол-3-фосфата) .
Далее фосфатидная кислотагидролизуется фосфатидат-фосфогидро-лазой до 1,2-диглицерида (1,2-диацилглицерола):
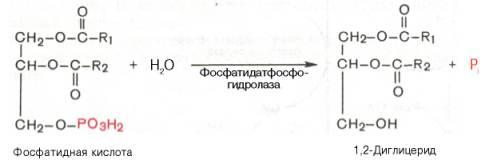
Затем 1,2-диглицерид ацилируется третьей молекулойацил-КоА и превращается в триглицерид (триацилглицерол). Этареакциякатализируется диацилглицерол-ацилтрансферазой:
