

53 Вопрос
Анаэробный гликолиз, гликолитическая оксидоредукция. Пируват как акцептор водорода; субстратное фосфорилирование. Ферменты гликолиза и «узкие звенья» гликолиза. Энергетическая эффективность. Роль в онтогенезе.
Анаэробный гликолиз– сложный ферментативный процесс распадаглюкозы, протекающий втканяхчеловека и животных без потреблениякислорода. Конечным продуктомгликолизаявляетсямолочная кислота. В процессегликолизаобразуетсяАТФ. Суммарное уравнениегликолизаможно представить следующим образом:
![]()
В анаэробных условиях гликолиз– единственный процесс в животноморганизме, поставляющий энергию. Именно благодарягликолизуорганизмчеловека и животных определенный период может осуществлять ряд физиологических функций в условиях недостаточностикислорода. В тех случаях, когдагликолизпротекает в присутствиикислорода, говорят об аэробномгликолизе.
гликолизпротекает в гиалоплазме (цитозоле)клетки.
Первой ферментативной реакциейгликолизаявляется фосфорилирование, т.е. перенос остаткаортофосфатанаглюкозуза счетАТФ.Реакциякатализируетсяферментомгексокиназой:
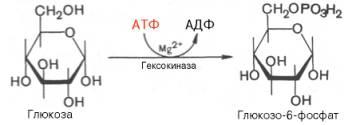
Образование глюкозо-6-фосфата в гексокиназной реакциисопровождается освобождением значительного количества свободной энергии системы и может считаться практически необратимым процессом.
Наиболее важным свойством гексокиназыявляется ее ингибирование глюкозо-6-фосфатом, т.е. последний служит одновременно и продуктомреакции, и аллостерическимингибитором.
Ферментгексокиназаспособен катализироватьфосфорилированиене только D-глюкозы, но и другихгексоз, в частности D-фруктозы, D-маннозы и т.д. Впечени, кромегексокиназы, существуетферментглюкокиназа, который катализируетфосфорилированиетолько D-глюкозы. Вмышечной тканиэтотферментотсутствует (подробнее см. главу 16).
Второй реакциейгликолизаявляется превращение глюкозо-6-фос-фата под действиемферментаглюкозо-6-фосфатизомеразы во фруктозо-6-фосфат:

Эта реакцияпротекает легко в обоих направлениях, и для нее не требуется каких-либо кофакторов.
Третья реакциякатализируетсяферментомфосфофруктокиназой; образовавшийся фруктозо-6-фосфат вновь фосфорилируется за счет второймолекулыАТФ:
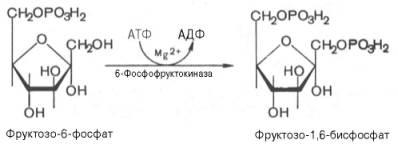
Данная реакцияаналогично гексокиназной практически необратима, протекает в присутствииионовмагнияи является наиболее медленно текущейреакциейгликолиза. Фактически этареакцияопределяет скоростьгликолизав целом.
Фосфофруктокиназа относится к числу аллостерических ферментов. Она ингибируетсяАТФи стимулируетсяАМФ. При значительных величинах отношенияАТФ/АМФактивностьфосфофруктокиназы угнетается игликолиззамедляется. Напротив, при снижении этого коэффициента интенсивностьгликолизаповышается. Так, в неработающей мышцеактивностьфосфофруктокиназы низкая, аконцентрацияАТФотносительно высокая. Во время работы мышцы происходит интенсивное потреблениеАТФиактивностьфосфофруктокиназы повышается, что приводит к усилению процессагликолиза.
Четвертую реакциюгликолизакатализируетферментальдолаза. Под влиянием этогоферментафруктозо-1,6-бисфосфат расщепляется на две фосфотриозы:
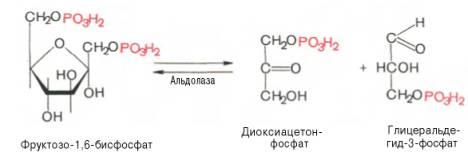
Эта реакцияобратима. В зависимости оттемпературыравновесиеустанавливается на различном уровне. При повышениитемпературыреакциясдвигается в сторону большего образования триозофосфатов (дигидро-ксиацетонфосфата и глицеральдегид-3-фосфата).
Пятая реакция– этореакцияизомеризациитриозофосфатов. Катализируетсяферментомтриозофосфатизомеразой:

Равновесиеданной изомеразнойреакциисдвинуто в сторону дигидроксиацетонфосфата: 95% дигидроксиацетонфосфата и около 5% глице-ральдегид-3-фосфата. В последующиереакциигликолизаможет непосредственно включаться только один из двух образующихся триозофосфатов, а именно глицеральдегид-3-фосфат. Вследствие этого по мере потребления в ходе дальнейших превращений альдегидной формы фосфотриозы ди-гидроксиацетонфосфат превращается в глицеральдегид-3-фосфат.
Образованием глицеральдегид-3-фосфата как бы завершается первая стадия гликолиза. Вторая стадия – наиболее сложная и важная. Она включаетокислительно-восстановительную реакцию(реакциягликолитической оксидоредукции), сопряженную с субстратнымфосфорилированием, в процессе которого образуетсяАТФ.
В результате шестой реакцииглицеральдегид-3-фосфат в присутствииферментаглицеральдегидфосфатдегидрогеназы,коферментаНАД инеорганического фосфатаподвергается своеобразномуокислениюс образованием 1,3-бисфосфоглицериновойкислотыи восстановленной формы НАД (НАДН). Этареакцияблокируется йод- или бромацетатом, протекает в несколько этапов:
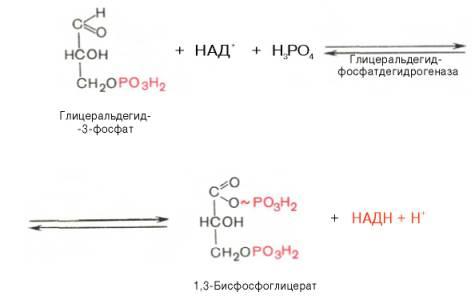
1,3-Бисфосфоглицерат представляет собой высокоэнергетическое соединение (макроэргическая связь условно обозначена знаком «тильда» ~). Механизм действия глицеральдегидфосфатдегидрогеназысводится к следующему: в присутствиинеорганического фосфатаНАД+ выступает как акцепторводорода, отщепляющегося от глицеральдегид-3-фосфата. В процессе образования НАДН глицеральдегид-3-фосфат связывается смолекулойферментаза счет SH-групп последнего. Образовавшаяся связь богата энергией, но она непрочная и расщепляется под влияниемнеорганического фосфата, при этом образуется 1,3-бисфосфоглицериноваякислота.
Седьмая реакциякатализируетсяфосфоглицераткиназой, при этом происходит передача богатого энергией фосфатного остатка (фосфатной группы в положении 1) наАДФс образованиемАТФи 3-фосфогли-цериновойкислоты(3-фосфоглицерат):
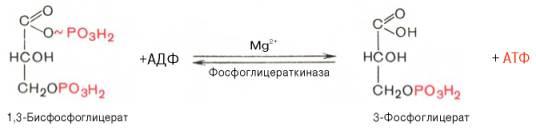
Таким образом, благодаря действию двух ферментов(глицеральде-гидфосфатдегидрогеназы ифосфоглицераткиназы) энергия, высвобождающаяся приокисленииальдегидной группы глицеральдегид-3-фосфата докарбоксильной группы, запасается в форме энергииАТФ. В отличие отокислительного фосфорилированияобразованиеАТФиз высокоэнергетических соединений называется субстратнымфосфорилированием.
Восьмая реакциясопровождается внутримолекулярным переносом оставшейся фосфатной группы, и 3-фосфоглицериноваякислотапревращается в 2-фосфоглицериновуюкислоту(2-фосфоглицерат).
Реакциялегкообратима, протекает в присутствииионовMg2+. Кофакторомферментаявляется также 2,3-бисфосфоглицериноваякислотааналогично тому, как в фосфоглюкомутазнойреакциироль кофактора выполняет глюкозо-1,6-бисфосфат:

Девятая реакциякатализируетсяферментоменолазой, при этом 2-фосфоглицериноваякислотав результате отщеплениямолекулыводыпереходит в фосфоенолпировинограднуюкислоту(фосфоенолпируват), а фосфатная связь в положении 2 становится высокоэргической:
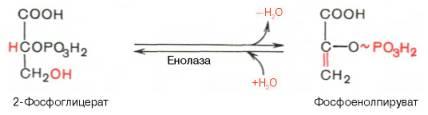
Енолазаактивируется двухвалентнымикатионамиMg2+или Мn2+ и ингибируетсяфторидом. Десятаяреакцияхарактеризуется разрывом высокоэргической связи и переносом фосфатного остатка от фосфоенолпирувата наАДФ(субстратноефосфорилирование). Катализируетсяферментомпируваткиназой:
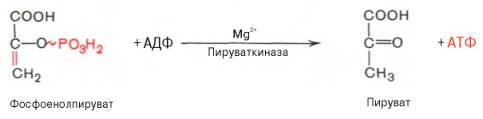
Для действия пируваткиназынеобходимыионыMg2+, а также одновалентныекатионыщелочных металлов(К+ или др.). Внутриклеткиреакцияявляется практически необратимой.
В результате
одиннадцатой реакциипроисходитвосстановлениепировиноградной
кислотыи образуетсямолочная
кислота.Реакцияпротекает
при участииферменталактатдегидрогеназыикоферментаНАДН,
образовавшегося в шестойреакции:
Реакциявосстановленияпирувата завершает внутренний окислительно-восстановительный циклгликолиза. НАД+ при этом играет роль промежуточного переносчикаводородаот глицеральдегид-3-фосфата (6-яреакция) напировиноградную кислоту(11-яреакция), при этом сам он регенерируется и вновь может участвовать в циклическом процессе, получившем название гликолитический оксидоредукции.
Биологическое значение процесса гликолизазаключается прежде всего в образовании богатых энергией фосфорных соединений. На первых стадияхгликолизазатрачиваются 2молекулыАТФ(гексокиназная и фосфофрук-токиназнаяреакции). На последующих образуются 4молекулыАТФ(фосфоглицераткиназная и пируваткиназнаяреакции). Таким образом, энергетическая эффективностьгликолизав анаэробных условиях составляет 2молекулыАТФна однумолекулуглюкозы. (На стадиях субстратного фосфорилирования образуется 4 молекулы АТФ, но две расходуются на начальных этапах гликолиза, так что "чистый выход" АТФ при гликолизе составляет 2 молекулы. В ходе расщепления Г-6-Ф, который образуется при распаде гликогена, энергетический выход несколько больше - 3 АТФ. Но, несмотря на малый выход АТФ, гликолиз очень важен, поскольку в отсутствие кислорода он является единствен¬ным энергодающим процессом в клетках.)
Как отмечалось, основной реакцией, лимитирующей скоростьгликолиза, является фосфофруктокиназная. Втораяреакция, лимитирующая скорость и регулирующаягликолиз– гексокиназнаяреакция. Кроме того, контрольгликолизаосуществляется также ЛДГ и ееизоферментами.