

17 Вопрос
Витамин С (аскорбиновая кислота). Структура, суточная потребность, пищевые источники, авитаминоз. Участие в окислительно-восстановительных процессах, стероидогенезе и образовании коллагена. Реакции гидроксилирования пролина и лизина.
Аскорбиновая
кислота - лактон кислоты, близкой по
структуре к глюкозе. Существует в двух
формах: восстановленной (АК) и окисленной
(дегидроаскорбиновой кислотой, ДАК).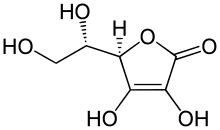
Обе эти формы аскорбиновой кислоты быстро и обратимо переходят друг в друга и в качестве коферментов участвуют в окислительно-восстановительных реакциях. Аскорбиновая кислота может окисляться кислородом воздуха, пероксидом и другими окислителями. ДАК легко восстанавливается цистеином, глутатионом, сероводородом. В слабощелочной среде происходят разрушение лактонового кольца и потеря биологической активности. При кулинарной обработке пищи в присутствии окислителей часть витамина С разрушается.
Источники витамина С - свежие фрукты, овощи, зелень, плоды шиповника, облепиха, смородина чёрная, лимоны, апельсины, яблоки.
Суточная потребность человека в витамине С составляет 50-75 мг.
Биологические функции. Главное свойство аскорбиновой кислоты - способность легко окисляться и восстанавливаться. Вместе с ДАК она образует в клетках окислительно-восстановительную пару с редокс-потенциалом +0,139 В. Благодаря этой способности аскорбиновая кислота участвует во многих реакциях гидроксилирования: остатков Про и Лиз при синтезе коллагена (основного белка соединительной ткани), при гидроксилировании дофамина, синтезе стероидных гормонов в коре надпочечников
В кишечнике аскорбиновая кислота восстанавливает Fe3+в Fe2+, способствуя его всасыванию, ускоряет освобождение железа из ферритина, способствует превращению фолата в коферментные формы. Аскорбиновую кислоту относят к природным антиоксидантам (см. раздел 8). Большое значение этой роли витамина С придавал известный американский учёный Л. Полинг, дважды лауреат Нобелевской премии. Он рекомендовал использовать для профилактики и лечения ряда заболеваний (например, простудных) большие дозы аскорбиновой кислоты (2-3 г).
Клинические проявления недостаточности витамина С. Недостаточность аскорбиновой кислоты приводит к заболеванию, называемому цингой (скорбут). Цинга, возникающая у человека при недостаточном содержании в пищевом рационе свежих фруктов и овощей, описана более 300 лет назад, со времени проведения длительных морских плаваний и северных экспедиций. Это заболевание связано с недостатком в пище витамина С. Болеют цингой только человек, приматы и морские свинки. Главные проявления авитаминоза обусловлены в основном нарушением образования коллагена в соединительной ткани. Вследствие этого наблюдают разрыхление дёсен, расшатывание зубов, нарушение целостности капилляров (сопровождающееся подкожными кровоизлияниями). Возникают отёки, боль в суставах, анемия. Анемия при цинге может быть связана с нарушением способности использовать запасы железа, а также с нарушениями метаболизма фолиевой кислоты.
18 Вопрос
Взаимосвязь обмена веществ и энергии. Экзергонические и эндергонические реакции в клетке. Типы макроэргических соединений (фосфатные, тиосульфатные). Строение АТФ, цикл АТФ/АДФ. Этапы унификации энергетических субстратов в организме: продукты, энергетическая ценность. Критические периоды развития ребенка и характеристика их обмена веществ.
Как было указано, обмен веществворганизмечеловека протекает не хаотично; он интегрирован и тонко настроен. Все превращения органическихвеществ, процессыанаболизмаикатаболизматесно связаны друг с другом. В частности, процессы синтеза и распада взаимосвязаны, координированы и регулируются нейрогормональными механизмами, придающими химическим процессам нужное направление. Ворганизмечеловека, как и в живой природе вообще, не существует самостоятельного обменабелков,жиров,углеводовинуклеиновых кислот. Все превращения объединены в целостный процессметаболизма. В настоящее время экспериментально обосновано существование четырех главных этапов распадамолекулуглеводов,белковижиров, которые интегрируют образование энергии из основных пищевых источников. На I этапеполисахаридырасщепляются домоносахаридов(обычногексоз);жирыраспадаются наглицеринивысшие жирные кислоты, абелки– на составляющие их свободныеаминокислоты. Следует подчеркнуть, что указанные процессы в основном являются гидролитическими, поэтому освобождающаяся в небольшом количестве энергия почти целиком используетсяорганизмамив качестве тепла.
На II этапе мономерные молекулы(гексозы,глицерин,жирные кислотыиаминокислоты) подвергаются дальнейшему распаду, в процессе которого образуются богатые энергией фосфатные соединения и ацетил-КоА. В частности, пригликолизегексозырасщепляются до пировиноград-нойкислотыи далее до ацетил-КоА. Этот процесс сопровождается образованием ограниченного числа богатых энергией фосфатных связей путем субстратногофосфорилирования. На этом этапевысшие жирные кислотыаналогично распадаются до ацетил-КоА, в то время какглицеринокисляется по гликолитическому пути допировиноградной кислотыи далее до ацетил-КоА. Дляаминокислотситуация на II этапе несколько отлична. При преимущественном использованииаминокислотв качестве источника энергии (при дефицитеуглеводовили присахарном диабете) некоторые из них непосредственно превращаются вметаболитылимоннокислого цикла (глутамат, аспартат), другие – опосредованно через глутамат(пролин,гистидин,аргинин), третьи – в пируват и далее в ацетил-КоА (аланин,серин,глицин,цистеин). Наконец, рядаминокислот, в частностилейцин, изо-лейцин, расщепляется до ацетил-КоА, а изфенилаланинаитирозина, помимо ацетил-КоА, образуется оксалоацетат черезфумаровую кислоту. Как видно, II этап можно назвать этапом образования ацетил-КоА, являющегося по существу единым (общим) промежуточным продуктомкатаболизмаосновных пищевыхвеществвклетках.
На III этапе ацетил-КоА (и некоторые другие метаболиты, например α-кетоглутарат, оксалоацетат) подвергаютсяокислению(«сгоранию») в цикле ди- и трикарбоновыхкислотКребса.Окислениесопровождается образованием восстановленных форм НАДН + Н+ и ФАДН2.
На IV этапе осуществляется перенос электроновот восстановленныхнуклеотидовнакислород(черездыхательную цепь). Он сопровождается образованием конечного продукта –молекулыводы. Этот транспортэлектроновсопряжен ссинтезом АТФв процессе окислительного фосфо-рилирования. 3. Эндергонические и экзергонические реакции
Направление химической реакции определяется значением AG. Если эта величина отрица-
тельна, то реакция протекает самопроизвольно и сопровождается уменьшением свободной энергии. Такие реакции называют экзергоничес-кими. Если при этом абсолютное значение AG велико, то реакция идёт практически до конца, и её можно рассматривать как необратимую.
Если AG положительно, то реакция будет протекать только при поступлении свободной энергии извне; такие реакции называют эн-дергоническими.
Если абсолютное значение AG велико, то система устойчива, и реакция в таком случае практически не осуществляется. При AG, равном нулю, система находится в равновесии (табл. 6-1).
4. Сопряжение экзергонических
и эндергонических процессов в организме
В биологических системах термодинамически невыгодные (эндергонические) реакции могут протекать лишь за счёт энергии экзер-гонических реакций. Такие реакции называют энергетически сопряжёнными. Многие из этих реакций происходят при участии аденозинтри-
|
|
Таблица 6-1. Соотношение между величинами K'eq и ΔG0' и направлением химических реакций

фосфата (АТФ), играющего роль сопрягающего фактора.
Рассмотрим подробнее энергетику сопряжённых реакций на примере фосфорилирования глюкозы.
Реакция фосфорилирования глюкозы свободным фосфатом с образованием глюкозо-6-фосфата является эндергонической:
(1) Глюкоза+ Н3РО4 → Глюкозо-6-фосфат + Н2О (ΔG = +13,8 кДж/моль).
Для протекания такой реакции в сторону образования глюкозо-6-фосфата необходимо её сопряжение с другой реакцией, величина свободной энергии которой больше, чем требуется для фосфорилирования глюкозы.
(2) АТФ → АДФ + Н3РО4 (ΔG = -30,5 кДж/моль).
При сопряжении процессов (1) и (2) в реакции, катализируемой гексокиназой (см. раздел 7), фосфорилирование глюкозы легко протекает в физиологических условиях; равновесие реакции
сильно сдвинуто вправо, и она практически необратима:
(3) Глюкоза + АТФ → Глюкозо-6-фосфат + АДФ (ΔG = -16,7 кДж/моль).
Макроэргические соединения – органические соединения живых клеток, содержащие богатые энергией, или макроэргические связи. Эти соединения образуются в результате фото- и хемосинтеза и биологического окисления. К ним относятся, например, вещества, при гидролизе которых высвобождается энергии в 2—4 раза больше, чем при гидролизе других веществ. К макроэргическим соединениям относятся аденозинтрифосфорная кислота (АТФ), аденозиндифосфорная кислота (АДФ), а также пирофосфат (H4P2O7), полифосфаты (полимеры метафосфорной кислоты — (НРО3)n * Н2О) и ряд других соединений. Самое важное макроэргическое соединение — АТФ. Используя энергию, заключенную в макроэргических связях АТФ, при действии ферментов, переносящих фосфатные группы, можно получить другие макроэргические соединения, например, ГТФ (гуанозинтрифосфорная кислота), ФЕП (фосфоенолпировиноградная кислота) и др. Образуется АТФ в процессах биологического окисления и при фотосинтезе. Аденозинтрифосфорная кислота (АТФ) - нуклеотид, образованный аденозином и тремя остатками фосфорной кислоты. Во всех живых организмах выполняет роль универсального аккумулятора и переносчика энергии. Под действием специальных ферментов концевые фосфатные группы отщепляются с освобождением энергии, которая идет на синтетические и другие процессы жизнедеятельности.
Аденозиндифосфат (АДФ) – нуклеотид, образованный аденозоном и двумя остатками фосфорной кислоты. Участвует в энергетическом обмене живых организмов. АДФ получает энергию путем дефосфорилирование фосфоэнолпировиноградной кислоты под действием фермента трансфосфорилазы, которая переносит макроэргическую связь с кислоты на АДФ. Уридиндифосфорная кислота (УДФ) и ее производные принимают участие во взаимопревращении углеводов. При биосинтезе гликозидной связи используется уридиндифосфатглюкоза (УДФГ), образующаяся из глюкозы‑1‑фосфата и уридинтрифосфата (УИФ). Если УДФГ передает глюкозу фруктозе, то образуется сахароза, а если цепочке декстрина – полисахарид. Аналогично образуются гликозиды, гликопротеиды и др. Взаимопревращение моносахаридов проходит через фосфорные эфиры сахаров или их уридиндифосфатпроизводные (УДФ-производные). УДФ-производные сахаров представляют собой тот или иной сахар, соединенный через два остатка фосфорной кислоты с уридином.
Сахарофосфаты являются источником фосфорного питания растений. Могут быть соли орто-, мета- и пирофосфорной кислоты и органические фосфаты. Лучшие из них – водорастворимые калиевые, натриевые, аммониевые, кальциевые и магниевые соли фосфорной кислоты.
Энергия макроэргических связей используется для совершения любой работы: активации соединений (например, глюкозы, чтобы могла начаться цепь ее окислительных превращений), синтеза биополимеров (нуклеиновых кислот, белков, полисахаридов), избирательного поглощения веществ из окружающей клетку среды и выброса из клетки ненужных продуктов, мышечного сокращения и восстановления активного состояния организма и т. д. Запас этих соединений позволяет организму быстро реагировать на изменение внешних условий и совершать физическую работу.
Цикл АТФ/АДФ.
АТФ - молекула, богатая энергией, поскольку она содержит две фосфоаншдридные связи (β, γ). При гидролизе концевой фосфоангидридной связи АТФ превращается в АДФ и ортофосфат Рi При этом изменение свободной энергии составляет -7,3 ккал/моль. При условиях, существующих в клетке в норме (рН 7,0, температура 37 °С), фактическое значение ΔG0' для процесса гидролиза составляет около -12 ккал/моль. Величина свободной энергии гидролиза АТФ делает возможным его образование из АДФ за счёт переноса фосфатного остатка от таких высокоэнергетических фосфатов, как, например, фосфоенолпируват
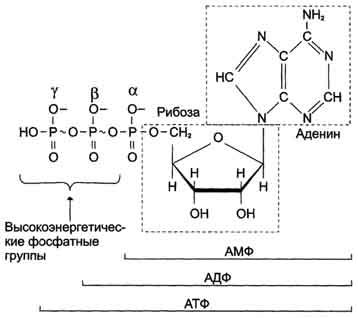
Рис. 6-2. Аденозинтрифосфорная кислота (АТФ). В молекуле АТФ две высокоэнергетические (макроэргические) связи β и γ, они обозначены на рисунке знаком ~ (тильда).
или 1,3-бисфосфоглицерат; в свою очередь, АТФ может участвовать в таких эндергонических реакциях, как фосфорилирование глюкозы или глицерина. АТФ выступает в роли донора энергии в эндергонических реакциях многих анаболических процессов. Некоторые биосинтетические реакции в организме могут протекать при участии других нуклеозидтрифосфатов, аналогов АТФ; к ним относят гуанозинтрифосфат (ГТФ), уридинтрифосфат (УТФ) и цитидинтрифосфат (ЦТФ). Все эти нуклеотиды, в свою очередь, образуются при использовании свободной энергии концевой фосфатной группы АТФ. Наконец, за счёт свободной энергии АТФ совершаются различные виды работы, лежащие в основе жизнедеятельности организма, например, такие как мышечное сокращение или активный транспорт веществ.
Таким образом, АТФ - главный, непосредственно используемый донор свободной энергии в биологических системах. В клетке молекула АТФ расходуется в течение одной минуты после её образования. У человека количество АТФ, равное массе тела, образуется и разрушается каждые 24 ч.
Использование АТФ как источника энергии возможно только при условии непрерывного синтеза АТФ из АДФ за счёт энергии окисления органических соединений (рис. 6-3). Цикл АТФ-АДФ - основной механизм обмена энергии в биологических системах, а АТФ - универсальная "энергетическая валюта".
Унификация энергетических субстратов в клетке Основными субстратами биоокисления являются углеводы, жиры и белки, весьма различные по своему составу. Филогенетически в организме животных выработалась система постепенной унификации (или стандарти¬зации) энергетических субстратов, повышающая эффективность окисле¬ния. Условно можно выделить два этапа унификации энергетического "то¬плива" в клетках. • На I этапе (переваривание в желудочно-кишечном тракте или распад в клетках) биополимеры расщепляются на свои структурные компоненты — мономеры, теряя при этом исходную специфичность строения. • На II этапе (тканевой обмен) мономеры в основном превращаются в пи-ровиноградную кислоту и/или далее в активную форму уксусной кисло¬ты - ацетил-КоА, который и является универсальным энергетическим субстратом. Затем в цикле трикарбоновых кислот Кребса происходит окисление (дегидрирование) ацетил-КоА с образованием восстановленных кофермен-тов НАД-Н и ФАД-Н2. В мембранах митохондрий они включаются в ды¬хательную цепь, где в ходе окислительного фосфорилирования в присутст¬вии кислорода происходит синтез АТФ из АДФ и фосфата. На I и II этапах унификации субстратов окисления выделяется до 40 %, в дальнейшем - около 60 % энергии. В связи с этим именно цикл трикарбоновых кислот считается основным "энергетическим котлом" клетки. При полном окислении до СОг и НгО одного грамма углеводов и белков образуется около 4,1 ккал, жира - 9,3 ккал энергии.