

Медленные тургорные настические движения
Рис. 13.9
Вьющийся побег рецкой фасоли
ту-
Обратимые тургорные движения осуществляются благодаря увеличению и уменьшению в вакуолях специализированных клеток концентрации осмотически активных веществ (К+, С1~, малата), в результате чего соответственно увеличиваются или уменьшаются поглощение воды и тургорное давление. Примерами могут служить медленные тургорные движения устьиц или опускание прекративших рост листьев на ночь («ночной сон»). Другой тип тургорных движений происходит за счет быстрого обратимого выделения жидкости из клеток в специали? зированных тканях (например, в сочленениях листьев мимозы).
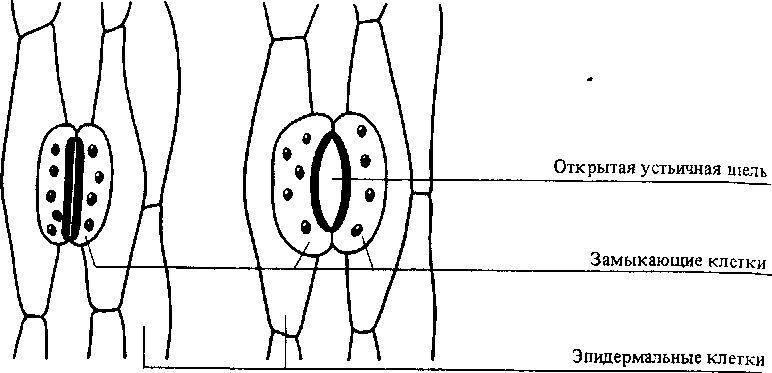
Движения устьиц. Примером обратимых движений являются движения устьиц. Эти движения обусловлены особенностью строения клеточных стенок замыкающих клеток. Утолщенной и нерастяжимой является та часть оболочки, которая обращена к устьичной щели. Остальная часть замыкающей клетки покрыта тонкой клеточной стенкой, способной эластически (обратимо) растягиваться (рис. 13.10). Такое строение приводит к тому, что при возрастании тургорного давления и увеличении объема вакуоли замыкающих клеток устьичные щели открываются (см. 5.4.3).
Степень их открытия определяется освещенностью, уровнем СОг в тканях и их оводненностью. Особенно чувствительны устьица к синим лучам. При восходе солнца устьица открываются при небольших интенсивностях света.
Открывание устьиц зависит от усиления выхода ионов Н+ из замыкающих клеток. Этот процесс связан с активацией Н+-помпы плазмалеммы в ответ на действие внешних или внутренних факторов. Освещение растения, адаптированного к темноте, приводит к усилению деятельности Н+-насосов плазмалеммы и тонопласта и гиперполяризации мембранного потенциала замыкающих клеток (рис. 13.11). Выход ионов Н+ из замыкающей клетки сопровождается одновременным проникновением ионов К+ в эту клетку и затем в ее вакуоль. Калий поступает из свободного пространства и из окружающих клеток. У галофитов функцию калия может выполнять натрий.
Транспорт К+в вакуоль происходит вместе с входом в нее анионов двух типов: ионов хлора, которые в зависимости от вида растения уравновешивают 5—100% поступившего К+, и анионов малата. Хлор транспортируется в замыкающие клетки из других клеток устьичного комплекса, а малат образуется в самих замыкающих клетках. Индуцированное светом усиление выхода Н+ из замыкающих клеток сопровождается некоторым подщелачиванием цитоплазмы (до рН 7,2 и выше), что увеличивает активность фосфоенолпируваткарбоксилазы (ФЕП-карбоксилазы), максимум активности которой наблюдается при рН 7,5 — 8,0. Фермент ускоряет процесс карбоксили-рования фосфоенолпирувата, образующегося из глюкозы при расщеплении крахмала, что приводит к возрастанию содержания малата в замыкающих клетках устьиц. Не исключена также возможность синтеза малата из гликолата в микротелах замыкающих клеток. Уровень малата в них регулируется отрицательной обратной связью между количеством образо-
Рис.
13.10
Устьица
в закрытом и открытом состоянии
-i20
К
„Ж1
в
3
вавшегося малата и активностью ФЕП-карбоксилазы, величиной рН цитоплазмы и наличием С02 в тканях. Именно необходимостью С02 для карбоксилирования ФЕП и образования малата объясняется чувствительность устьиц к изменениям концентрации С02 в тканях.
Увеличение в вакуолях замыкающих клеток содержания осмотически активных веществ (К + , малата, С1~) приводит к усилению поступления воды в вакуоли, к возрастанию тур-горного давления, обратимому растяжению эластичных участков клеточных стенок и открыванию устьичной щели. В регуляции открывания устьиц могут участвовать и цитокинины, способные усиливать поглощение К+ замыкающими клетками устьиц, активируя, возможно, Н+,К + -АТРазу плазмалеммы.
При закрывании устьиц в замыкающих клетках содержание К + , С1~ и малата уменьшается. Этот процесс регулируется С02 и абсцизовой кислотой. При высоких концентрациях ССЬ внутриклеточная среда в замыкающих клетках закисляется, что снижает активность ФЕП-карбоксилазы. АБК, содержание которой возрастает при водном дефиците (см. 5.4.3), вызывает закрывание устьиц, нарушая Н+/К+ обмен в замыкающих клетках (гигронастия).
Таким образом, процесс открывания и закрывания устьиц связан с транспортом ионов через плазмалемму и тонопласт замыкающих клеток и изменением в них тургорного давления. Сходные механизмы лежат в основе всех тургорных настиче-ских движений органов растений в ответ на смену диффузно действующих факторов внешней среды.
Тургорные фотонастии. Смена условий освещения в течение суток вызывает медленные ритмические движения листьев многих бобовых (фасоли, клевера и т. д.) и ряда растений других семейств (например, кислицы). Эти движения называют ник-тинастическими, или движениями «сна».
Движения листьев обеспечиваются деятельностью специализированных образований: подушечек в сочленениях между стеблем и черешком, между черешком и пластинкой листа. У мимозы стыдливой имеются гри порядка таких сочленений: между стеблем и главным черешком (подушечки первого порядка), между главным и боковыми черешками (второго порядка) и в основаниях мелких листовых пластинок (третьего
Вакуоль
Механизм
осморегуля-ции движений замыкающих
клеток устьиц с участием Н +
-помпы (по К. Raschke,
1976)

порядка). Функцию моторных клеток в подушечках выполняют паренхимные клетки коры. Если моторные клетки при складывании листьев способны терять тургор и восстанавливать его при открывании, то их называют экстензорными (разгибателями).
Клетки, тургор которых возрастает при складывании листьев и снижается при их открывании, называют флексорны-ми (сгибателями). Клеточные стенки экстензорных клеток эластичнее, чем у флексорных. Если экстензоры находятся на верхней стороне подушечки (например, у третичных подушечек стыдливой мимозы и акации шелковой), то листочки на ночь складываются вверх. Если экстензорные клетки расположены снизу подушечки, то листья складываются вниз (например, вторичные подушечки фасоли, кассии).
Никтинастические движения листьев связаны с изменениями транспорта- ионов и тургора в моторных клетках. При этом моторные клетки взаимодействуют с другими клетками коры и клетками колленхимы подушечки. Как и в устьичных клетках, изменения ионных потоков при смене режима освещения связаны с деятельностью Н + -помп, выводящих Н+ из моторных клеток, с транспортом К+, С1~ и других анионов. При движении взаимодействуют Н+-помпы моторных клеток верхних и нижних частей подушечки, работающие в противоположных фазах (рис. 13.12). Складывание листочков с наступлением темноты обусловлено снижением активности Н+-помпы экстензорных клеток, что вызывает выход из вакуоли этих клеток К и О", а вслед за ними воды в утолщенные клеточные стенки колленхимы и в флексорные клетки, где активность Н+-помпы продолжает оставаться высокой. Это поддерживает поступление в них К + , анионов и Сахаров с помощью Н+-ко-транспортного механизма. Уменьшение тургора в экстензорных клетках приводит к складыванию листьев. Открывание листьев вызывается активацией Н +-помпы в экстензорных клетках, увеличением содержания в них К+, С1~, Сахаров, повышением тургора и объема клеток. При этом во флексорных клетках тургор снижается.
Никтинастические движения могут регулироваться фитогор-монами (например, ауксином), влияющими на деятельность Н+-помпы. Обработка листьев этиленом приводит к хорошо выраженной эпинастии (их опусканию). Движения чувстви-

МК — моторная клетка, Клх — колленхима, ПК — переходная клетка. Широкая стрелка означает максимальную активность Н+-насоса, тонкая — минимальную. Заштрихованы суберинизи-рованные клеточные стенки
тельны к действию температуры, причем ее снижение ускоряет закрывание листьев и замедляет открывание (тер-монастии).
Тигмонастии. Железистые волоски росянки отвечают на прикосновение шершавых предметов изгибанием и выделением муравьиной кислоты. На капли дождя росянка не реагирует. Чувствительность к механическому раздражению (трению) очень высока. Реакция начинается от действия отрезка волоска (нитки) массой 0,82 мкг. Раздражение воспринимается самым кончиком железистого волоска. Затем раздражение передается по цитоплазме нижележащим клеткам. Визуально это выражается в сокращении («агрегации», как назвал Дарвин) вакуолей. Скорость проведения 8 мм/мин. Волосок изгибается ближе к основанию, т. е. место рецепции отделено от места реакции. Время от момента раздражения до начала реакции у росянки составляет около 1 мин. В настоящее время известно, что стимуляция механорецепторов головок волосков росянки вызывает появление нераспространяющихся рецепторных потенциалов в соответствии с силой и продолжительностью раздражения. Если продолжительность и амплитуда рецепторных потенциалов достаточно велики, то индуцируется возникновение распространяющихся потенциалов действия. Так обеспечивается передача раздражения к месту реакции.
Хемонастии. Изгибы на химическое раздражение у росянки выражены даже в большей степени, чем на механическое. Хе-морецепторной функцией обладает также головка волоска (рис. 13.13). Результатом хеморецепции служит индукция сгибания волоска и выделения пищеварительного сока. Хеморецепторы росянки наиболее чувствительны к азотсодержащим веществам
Рис.
13.13 Лист росянки. А — невоэбуж-денный
лист; Б — частичная реакция листа
на раздражение; В — головка железистого
волоска

13.6.2