

Но
N-
пировиноградная
кислотаГлутаминовая кислота

(1-й
<' И -NH, {'ООН
Трансаминирование с использованием аминогруппы глутамата широко распространено в растениях, обеспечивая возможность синтеза ряда аминокислот. Наиболее важные акцепторы NH2-rpynnbi в процессе трансаминирования и соответствующие им аминокислоты приведены ниже:
Акцепторы ^Ш2-группы
а - Кетоглутаровая
Щавелевоуксусная
Глиоксиловая
Пировиноградная
Гидроксипировиноградная
Фенилпировиноградная
Аминокислоты
Глутаминовая
Аспарагиновая
Глицин
Аланин
Серии
Фенилаланин
ch. on
( 11 Nil (ООП
При обильном снабжении растений аммонийными источниками азота в их тканях в значительном количестве накапливаются амиды — глутамин и аспарагин. Это явление было обнаружено и изучалось в классических работах Э. Шульце, Д. Н. Прянишникова, А. Чибнелла, а в последние 30 лет -
СООН
I
СН2
I
ch-nh,
СООН
Аепарапшовая кислот а
со - \н
сн,
I -
СН N 11 СООН
В. Л. Кретовичем с сотрудниками. Особенно большой вклад в изучение роли амидов в растениях внес Прянишников, который рассматривал эти соединения как обезвреживающую, запасающую и транспортную форму азота в растениях. Исследования Кретовича с сотрудниками показали, что 15N включается в амидные группы амидов в 2 — 3 раза интенсивнее, чем в амин-ные. В то же время у большинства растений глутамин преобладает над аспарагином.
Синтез глутамина был рассмотрен в разделе 6.3.4. Синтез аспарагина у растений может происходить двумя путями:
\ Mg2 +
Аспарагиновая кислота (L-аспартат) + NH3 + АТР —> L-Ac-парагин + ADP + Р,
L-Аспартат + L-глутамин + АТР -»L-Аспарагин + Глутамат + AMP + РР,
Обе реакции катализируются ферментом аспарагинсинтетазой. Первая реакция прямого синтеза аспарагина из аспараги-новой кислоты и NH3 аналогична синтезу глутамина, катализируемому глутаминсинтетазой. Во второй реакции аспа-рагин образуется в результате переаминирования глутамина и аспарагиновой кислоты. Механизмы этих реакций пока мало исследованы. Аспарагинсинтетаза локализована в цитоплазме. Второй путь синтеза аспарагина, при котором донором NH2-rpynnbi служит глутамин, по-видимому, значительно более распространен в растениях, так как сродство фермента ас-парагинсинтетазы к глутамину в 10 раз выше, чем к иону NH4+. Аспарагиновая кислота возникает в результате переаминирования с участием щавелевоуксусной кислоты. Прямой синтез аспарагина в соответствии с первой реакцией может осуществляться в условиях избыточно высокого уровня NH^ в тканях.
При выращивании растений на аммонийных источниках азота связывание NH^ в виде амидов происходит уже в корнях. В этом случае более половины азота, транспортируемого из корней с пасокой, находится в форме амидов. Участие корней в связывании поглощенного аммония подтверждает представление Д. А. Сабинина, развитое в дальнейшем акад. А. Л. Курсановым, об активной роли корневой системы в синтезе органических соединений (рис. 6.4.). В корнях в результате процессов аминирования, амидирования и переаминирования могут синтезироваться более 25 видов азотных соединений. Роль углеродных скелетов в этих реакциях играют органические кислоты—продукты гликолиза и цикла Кребса. Практически весь азот, поглощенный в форме NH% ассимилируется в корневой системе и поступает в надземные части в виде аминокислот и амидов.
В условиях нитратного питания ассимиляция NOf у растений осуществляется и в листьях. В этом случае акцепторами выступают первичные продукты фотосинтез* и фотодыхания. Поэтому набор аминокислот, синтезируемых в листьях, может быть качественно иным: больше синтезируется сложных и ароматических аминокислот.
Рис.
6.4
Круговорот
веществ в целом растении (по А. Л.
Курсанову, 1976)
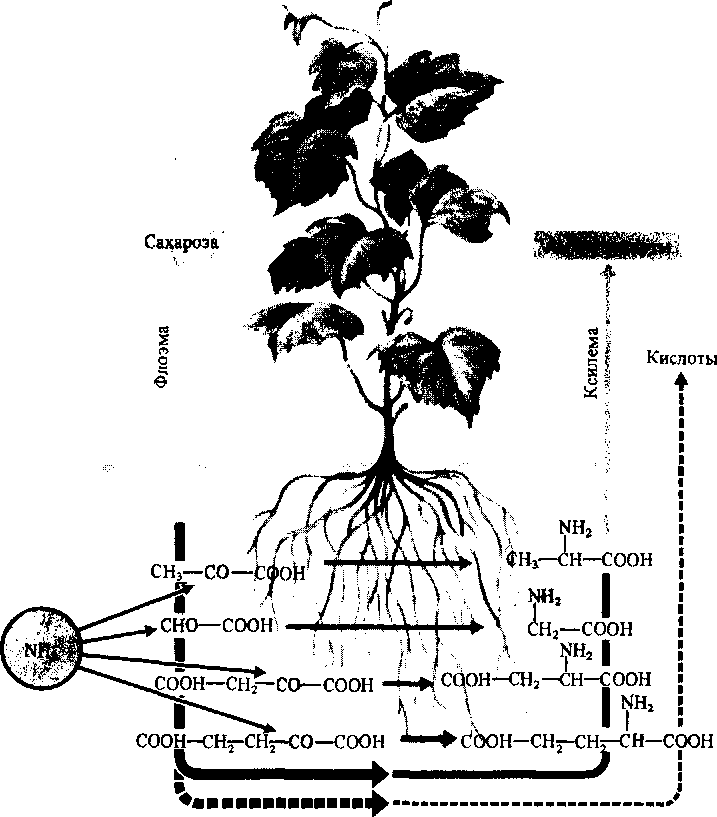
Аминокислоты
Амиды Азотистые основания

Нуклеиновые кислоты