

3.6. Экология фотосинтеза
111
Спектральный
состав прямой солнечной радиации
при разной высоте стояния солнца
над горизонтом (по Н. Н. Калитину, 1947)

лизме углерода на синем свету у зеленых растений, являются флавины. Скорость фотосинтеза быстро и значительно увеличивается при добавке небольшого количества (20% от насыщения красного света) синего света к красному. По-видимому, это связано с тем, что фотохимическая стадия фотосинтеза регулируется синим светом.
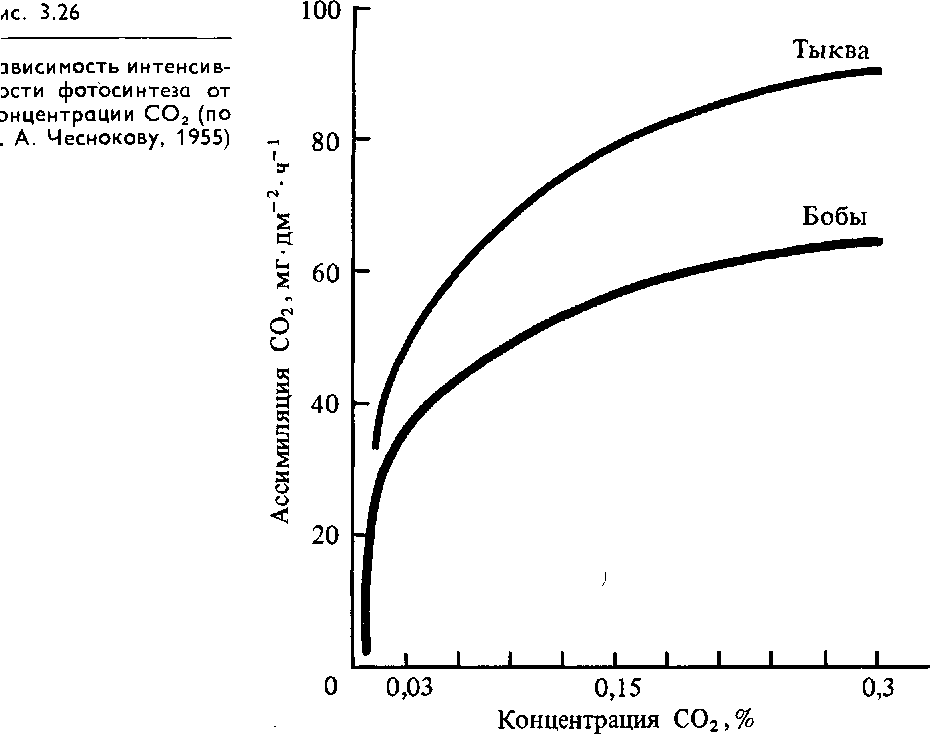
Концентрация диоксида углерода. С02 является основным субстратом фотосинтеза; его содержание определяет интенсивность процесса. Концентрация С02 в атмосфере составляет 0,03%. В слое воздуха высотой 100 м над гектаром пашни содержится 550 кг С02. Из этого количества за сутки растения поглощают 120 кг С02. Зависимость фотосинтеза от С02 выражается логарифмической кривой (рис. 3.26). При концентрации 0,03 % интенсивность фотосинтеза составляет лишь 50% от максимальной, которая достигается при 0,3% С02. Это свидетельствует о том, что в эволюции процесс фотосинтеза формировался при большей концентрации С02 в атмосфере. Кроме того, такой ход зависимости продуктивности фотосинтеза от концентрации С02 указывает на возможность подкормки растений в закрытых помещениях С02 для получения большего урожая. Такая подкормка С02 оказывает сильное влияние на урожай растений с Сз-типом ассимиляции С02 и не влияет на растения с С4-типом, у которых существует особый механизм концентрирования С02 (см. рис. 3.14).
Интенсивность ассимиляции С02 зависит от скорости его поступления из атмосферы в хлоропласты, которая определяется скоростью диффузии С02 через устьица, межклетники и в цитоплазме клеток мезофилла листа. В открытом состоянии устьица занимают лишь 1—2% площади листа, остальная поверхность покрыта плохо проницаемой для газов кутикулой. Однако при наличии кутикулы С02 входит в лист через устьица за единицу времени в таком же количестве, как и без нее. Объясняется это законом Стефана, согласно которому скорость перемещения молекул газа через малые отверстия пропорциональна их окружности, а не площади. Чем меньше отверстие, тем больше отношение окружности к площади. А у края отверстия молекулы в меньшей степени сталкиваются друг с другом и быстрее диффундируют. Поэтому через устьице с апертурой (открытостью)
порядка 10 мкм молекулы газа перемещаются с большой скоростью. На процессы открывания и закрывания устьиц влияют С02, насыщенность тканей водой, свет, фитогормоны (см. 13.6.1).
Температура. Первичные фотофизические процессы фотосинтеза (поглощение и миграция энергии, возбужденные состояния) не зависят от температуры. Очень чувствительны к температуре процессы фотосинтетического фосфо12шгщиг]шщя. Скорость комплекса энзиматических реакций, сопряженных с восстановлением углерода, при повышении температуры на 10°С возрастает в 2-3 раза (Q10 = 2-3).
Общая зависимость фотосинтеза от температуры выражается одновершинной кривой (рис. 3.27). Кривая имеет три основные (кардинальные) температурные точки: минимальную, при которой начинается фотосинтез, оптимальную и максимальную. Интенсивность фотосинтеза при супероптимальных температурах зависит от продолжительности их воздействия на растения. Нижняя температурная граница фотосинтеза у растений северных широт находится в пределах — 15 °С (сосна, ель)... —0,5 °С, а у тропических растений — в зоне низких положительных температур 4 — 8 °С. У растений умеренного пояса в интервале 20 — 25 °С достигается максимальная интенсивность фотосинтеза, а дальнейшее повышение температуры до 40 °С приводит к быстрому ингибированию процесса (при 45 °С растения погибают). Некоторые растения пустынь способны осуществлять фотосинтез при 58 °С. Температурные границы
Зависимость
интенсивности фотосинтеза у ели от
температуры (Б. А. Рубин, В. Ф. Гав-риленко,
1977)
О 10 20 30 40
Т,°С
фотосинтеза можно раздвинуть предварительным закаливанием, адаптацией растений к градиенту температур. Наиболее чувствительны к действию температуры реакции кар-боксилирования, превращения фруктозо-6-фосфата в сахарозу и крахмал, а также транспорт сахарозы из листьев в другие органы.
Необходимо отметить, что влияние на фотосинтез света, концентрации С02 и температуры осуществляется в сложном взаимодействии. Особенно тесно взаимосвязаны свет, действующий на скорость фотохимических реакций, и температура, контролирующая скорость энзиматических реакций. В условиях высокой интенсивности света и низких температур (5—10 °С), когда главным фактором, лимитирующим скорость всего процесса, являются ферментативные реакции, контролируемые температурой, значения Q10 могут быть > 4. При более высоких температурах Q10 снижается до 2. При низких ин-тенсивностях света Q10 = 1, т. е. фотосинтез относительно независим от температуры, так как его скорость в данном случае ограничивается фотохимическими реакциями.
Водный режим. Вода непосредственно участвует в фотосинтезе как субстрат окисления и источник кислорода. Другой аспект влияния содержания воды на фотосинтез состоит в том, что величина оводненности листьев определяет степень открывания устьиц и, следовательно, поступления С02 в лист. При полном насыщении листа водой устьица закрываются, что снижает интенсивность фотосинтеза. В условиях засухи чрезмерная потеря воды листом также вызывает закрывание устьиц под влиянием увеличения содержания в листьях абсцизовой кислоты в ответ на недостаток влаги. Длительный водный дефицит в тканях листа при засухе приводит к ингибированию нециклического и циклического транспорта электронов и фотофосфорилирования и к снижению величины отношения ATP/NADPH за счет большего тормо-l жения образования АТР. Максимальный фотосинтез наблюдается при небольшом водном дефиците листа (порядка 5 — 20% от полного насыщения) при открытых устьицах.
Минеральное питание. Для нормального функционирования фотосинтетического аппарата растение должно быть обеспечено всем комплексом макро- и микроэлементов. Два основных процесса питания растительного организма — воздушный и корневой — тесно взаимосвязаны. Зависимость фотосинтеза от элементов минерального питания определяется их необходимостью для формирования фотосинтетического аппарата (пигментов, компонентов электронтранспортной цепи, каталитических систем хлоропластов, структурных и транспортных белков), а также для его обновления и функционирования. (_Магний входит в состав хлорофиллов, участвует в деятельности сопрягающих белков при синтезе АТР, влияет на активность реакций карбоксилирования и восстановления NADP+. Вследствие этого его недостаток нарушает процесс фотосинтеза.
Железо в восстановленной форме необходимо для процессов биосинтеза хлорофилла и железосодержащих соединений хлоропластов (цитохромов, ферредоксина). Дефицит железа резко нарушает функционирование циклического и нециклического фотофосфорилирования, синтез пигментов и изменяет структуру хлоропластов.
Необходимость марганца для зеленых растений связана с его ролью в фотоокислении воды. Поэтому недостаточность питания по марганцу отрицательно сказывается на интенсивности фотосинтеза. В реакциях фотоокисления воды необходим также хлор.
Медь входит в состав пластоцианина, поэтому у растений дефицит меди вызывает снижение интенсивности фотосинтеза.
Недостаток азота сильно сказывается на формировании пигментных систем, структур хлоропласта и его общей активности. Концентрация азота определяет количество и активность РДФ-карбоксилазы.
В условиях недостатка фосфора нарушаются фотохимические и темновые реакции фотосинтеза. Особенно резко дефицит фосфора проявляется при высокой интенсивности света, при этом более чувствительными оказываются темновые реакции. Однако при уменьшении содержания фосфора в два раза интенсивность фотосинтеза снижается в меньшей степени, чем ростовые процессы и общая продуктивность растений. Избыток фосфора также тормозит скорость фотосинтеза, по-видимому, вследствие изменения проницаемости мембран.
Уменьшение содержания калия в тканях сопровождается значительным снижением интенсивности фотосинтеза и нарушениями других процессов в растении. В хлоропластах раз-