

Структуры растительных клеток
белка, что может приводить к слипанию белков-ферментов и нарушению их функций. Поэтому для деятельности мембранных ферментов важно, чтобы окружающие их липиды находились в жидком агрегатном состоянии.
Непроницаемость фосфолипидного бислоя для большинства соединений создает условия для векторных процессов. Векторный характер имеют транспорт веществ и передача сигналов из внешней среды в цитоплазму. Само расположение ферментов в липидной фазе мембраны весьма существенно для их активности. Таким образом, благодаря упорядоченности и векторности ферментов и транспортных механизмов осуществляется мембранная координация множества биохимических реакций, протекающих в клетке.
Аккумуляция и трансформация энергии — одна из важнейших функций мембран. В хлоропластах зеленых растений энергия света трансформируется ~в~знергию NADH и АТР, а в конечном счете — в стабильную энергию химических связей Сахаров, органических кислот, аминокислот. Эти ассимиляты, окисляясь затем в цитоплазме, и митохондриях, освобождают энергию, необходимую клеткам для выполнения различных функций. Согласно хемиосмоти-ческой теории П. Митчелла, процесс синтеза АТР в митохондриях и хлоропластах опосредован образованием электрохимического мембранного потенциала ионов Н+ (см. рис. 1.3) за счет работы Н +-насоса.
Помимо редокс-цепей и Н + -АТРазы в клетках имеются и другие насосы, создающие электрохимические потенциалы: Na+, К + -АТРаза, Са2+-АТРаза, анионная АТРаза. Как отмечает В. П. Скулачев (1978), осмотический градиент этих потенциалов — трансмембранный градиент К+ (АрК + ) и Na+ (ApNa+) — нужно рассматривать как резервную форму энергии. Таким образом, в живых клетках помимо дыхательных субстратов энергия может запасаться в двух формах — в виде легко отдающих энергию химических соединений (АТР, NADH и др.) и в виде электрохимического мембранного потенциала (ДДн+> ДДк+> A^Na+)- Все эти резервные формы энергии способны к взаимопревращениям, а также могут быть использованы на другие виды работ: химическую, осмотическую, тепловую и т. д.
Важнейшая функция мембран — рецепторн о-регуляторная. В состав мембран входят хемо-, фото- и механоредепторы белковой природы, чувствительные к действию химических и физических факторов. Эти рецепторы воспринимают сигналы, поступающие из внешней и внутренней среды, обеспечивая адаптивные (приспособительные) ответы клеток на изменения условий существования.
Ядро. В растительной клетке ядро имеет диаметр порядка 10 мкм. Оно может быть сферическим, удлиненным или лопастным. Внутреннее содержимое ядра (нуклеоплазма) ограничено ядерной оболочкой, состоящей из двух элементарных мембран — внутренней и наружной. Ядерная оболочка пронизана порами диаметром 10 — 20 нм, через которые
транспортируются нуклеиновые кислоты и белки. С наружной мембраной ядерной оболочки могут быть непосредственно связаны элементы эндоплазматического ретикулума. В нуклео-плазме локализованы участки хроматина, состоящего из комплекса ДНК, РНК и белков. В делящихся клетках хроматин организуется в хромосомы, число которых специфично для каждого вида растений. Количество ДНК в одном ядре постоянно для каждого вида (кроме случаев мейоза и полиплоидии). Геном растений отличается от генома животных болящим количеством ДНК. В нуклеоплазмё ядра содержатся также ферменты и кофакторы, необходимые для обеспечения процессов репликации и транскрипции ДНК, молекулы различных РНК, ферменты, фосфорилирующие и ацетилирую-щие ядерные белки, ферменты гликолиза и др.
Ядрышко отчетливо видно в ядре под световым и электронным микроскопами. Оно формируется на определенных участках ДНК, называемых ядрышковым организатором. В хроматине ядрышка находятся участки ДНК, ответственные за синтез рибосомальных РНК (рРНК).
В целом ядро является местом хранения генетической информации клетки и репликации ДНК. В нем происходит процесс транскрипции ДНК в РНК различных типов. В тесном взаимодействии с цитоплазмой ядро участвует в обеспечении экспрессии генетической информации и контролирует процессы жизнедеятельности клетки.
Рибосомы. Рибосомы осуществляют синтез белков — трансляцию матричной, или информационной, РНК (мРНК).'~ На электронных фотографиях они выглядят округлыми частицами диаметром 20 — 30 нм. Каждая рибосома состоит из двух нуклеопротеиновых субъединиц. В цитоплазме растительных клеток находятся, 80 S рибосомы, состоящие из 40 и 60 S субъединиц, в хлоропластах — 70 S Джбосомы, а в митохондриях — 78 — 80 S рибосомы, отличные от цитоплазмати-ческих и хлоропластных. Субъединицы рибосом, образованные в ядрышке, поступают в цитоплазму, где происходит сборка рибосом на молекуле мРНК.
Рибосомы в цитоплазме могут быть свободными, прикрепленными к мембранам эндоплазматического ретикулума, к наружной мембране ядерной оболочки либо образуют поли-рибосомные (полисомные) комплексы. Полирибосомы возникают потому, что одну молекулу мРНК могут одновременно транслировать несколько рибосом. Полирибосомы разрушаются при воздействии на растения неблагоприятных факторов внешней среды (например, засухи, недостатка кислорода). В процессе синтеза белка, осуществляемого рибосомами, участвуют компоненты, образованные в ядре, ядрышке, в цитоплазме, митохондриях и хлоропластах.
Пластиды. Для клеток растений типична система органоидов, называемая пластидной. Пластиды — это ограниченные двойной мембраной округлые или овальные органоиды, содержащие внутреннюю систему мембран. Пластиды могут быть бесцветными (пропластиды, лейкопласты, этиоплас-
ты) или окрашенными (хлоропласты, хромопласты) (см. рис. 10.4).
В меристемах присутствуют пропластиды, у которых внутренняя мембрана имеет лишь небольшие инвагинации (впячивания). Если структура пропластид сохраняется у органоидов зрелых клеток, их называют лейкопластами. В лейкопластах откладываются запасные вещества и названия они получают в зависимости от этих соединений: если запасается крахмал — амилопласты, жиры — элайопласты, белки — про-теинопласты и т. д. Этиопласты формируются при выращивании зеленых растений в темноте. При освещении они превращаются в хлоропласты, при этом внутренняя система мембран подвергается сложным перестройкам (см. рис. 10.5).
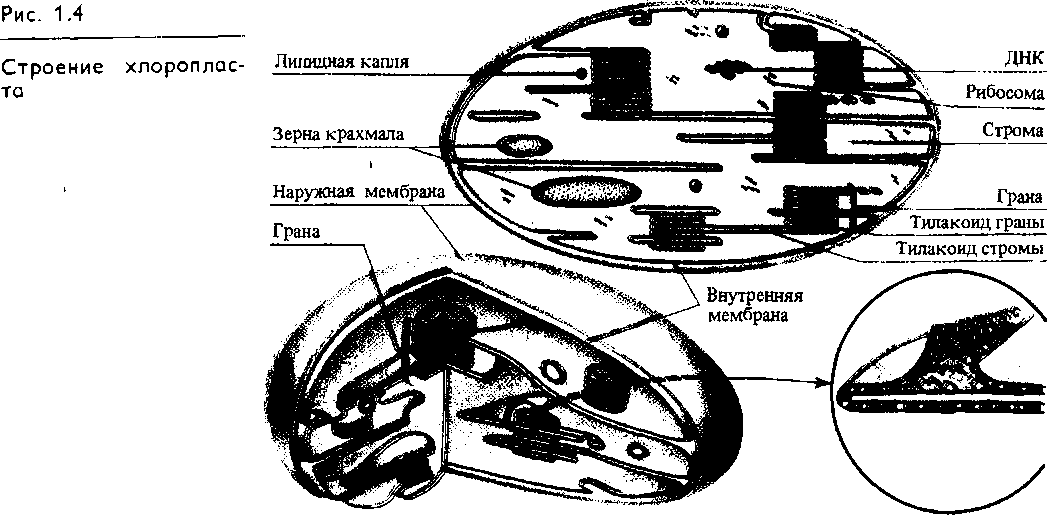
Важнейший для всего живого процесс преобразования энергии поглощенного света в химическую энергию органических веществ, синтезируемых из С02 и Н20 (фотосинтез), осуществляется в зеленых пластидах — хлоропластах. Это овальные тела 5—10 мкм длиной и 2 — 3 мкм в диаметре. В одной клетке листа могут находиться 15 — 20 и более хлоропластов, а у некоторых водорослей — лишь 1 — 2 гигантских хлоропласта различной формы. Как и другие пластиды, хлоропласты обладают наружной и внутренней мембранами (рис. 1.4). Внутренняя мембрана, ограничивающая внутреннюю гомогенную среду (строму) хлоропласта, образует уплощенные инвагинации — тилакоиды, которые могут иметь форму дисков и в этом случае называются тилакоидами гран (гранальными). Несколько таких тилакоидов, лежащих друг над другом, формируют стопку — грану. Другие тилакоиды, связывающие между собой граны или не контактирующие с ними, называются тилакоидами стромы.
В мембранах тилакоидов локализованы зеленые (хлоро-филлы), желтые и красные (каротиноиды) пигменты, компоненты редокс-цепей и запасания энергии, участвующие в погло-

щении и использовании энергии света. Биохимические системы синтеза и превращения углеводов функционируют в строме хлоропластов. В ней же может откладываться крахмал.
Хромопласты содержат в везикулах (пузырьках) стромы каротиноиды. Их присутствием объясняется окраска плодов томатов, рябины и др. В строме всех пластид обнаружены кольцевые молекулы ДНК.
При росте клетки количество хлоропластов увеличивается путем деления. При делении ламеллярная система пересекается перемычкой поперек органоида, иногда наблюдается почкование хлоропластов. Затем размер дочерних хлоропластов увеличивается. Деление хлоропластов происходит через 6 —20 ч и не обязательно совпадает с делением ядра. Оно может регулироваться красным светом (660 нм) и устраняется облучением дальним красным светом (730 нм). Деление останавливается также низкой температурой.
Митохондрии. В растительных клетках митохондрии представлены округлыми или гантелевидными Делами диаметром 0,4 — 0,5 мкм и длиной 1 — 5 мкм. Количество митохондрий варьирует от десятков до 2000 на клетку, меняясь в онтогенезе клетки и в зависимости от ее функционального состояния. Органоиды ограничены наружной и внутренней мембранами толщиной 5 — 6 нм каждая (рис. 1.5). Внутренняя мембрана образует складки различной формы, называемые кристами (гребнями). Мембраны отделяют от цитоплазмы внутреннее содержимое митохондрий — матрикс. В матрик-се содержатся рибосомы и митохондриальная ДНК, ответственные за синтез части белков митохондрий. ДНК в митохондриях имеет кольцевое строение.
Собственная генетическая система и способность мито-хондриальной ДНК к репликации позволяют митохондриям делиться независимо от деления ядра. В клетке они образуются от предсуществующих митохондрий. Продолжительность жизни (оборот) митохондрий определяется скоординированной деятельностью ядра, цитоплазмы и самих митохондрий. Полупериод жизни этих органоидов у различных растений составляет 5—10 дней, причем наружная мембрана обновляется быстрее внутренней.


 В
митохондриях^функционируют системы
аэробного .дыхания и
окислителънохофесфэршгитда'вания,
обеспечивающие энергетические,
потребности клеток. Во внутренней
мембране митохондрий локализованы
компоненты электронтранспортной цепи
и АТР-синтетазные комплексы, осуществляющие
транспорт электронов и протонов и
сопряженный с ним синтез АТР. В матриксе
располагаются ферментные системы
окисления ди- и трикарбоновых кислот,
а также ряд систем синтеза липидов,
аминокислот и др.
В
митохондриях^функционируют системы
аэробного .дыхания и
окислителънохофесфэршгитда'вания,
обеспечивающие энергетические,
потребности клеток. Во внутренней
мембране митохондрий локализованы
компоненты электронтранспортной цепи
и АТР-синтетазные комплексы, осуществляющие
транспорт электронов и протонов и
сопряженный с ним синтез АТР. В матриксе
располагаются ферментные системы
окисления ди- и трикарбоновых кислот,
а также ряд систем синтеза липидов,
аминокислот и др.
Перокснсомы н глнокснсомы. В растениях присутствуют округлые органоиды диаметром 0,2—1,5 мкм, ограниченные элементарной мембраной и содержащие гранулярный матрикс умеренной электронной плотности. Они получили название микротел. В некоторых микротелах обнаруживается белковый кристаллоид, состоящий из трубочек диаметром около 6 нм. Количество микротел в клетке близко к числу митохондрий. В клетках растений обнаружены два типа микротел, выполняющих различные физиологические функции: пероксисомы и глиоксисомы.
Пероксисомы многочисленны в клетках листьев, где они тесно связаны с хлоропластами. В них окисляется синтезируемая в хлоропластах в ходе фотосинтеза гликолевая кислота и образуется аминокислота глицин, которая в митохондриях превращается в серии. В листьях высших растений пероксисомы участвуют в фотодыхании (см. рис. 3.16).
Глиоксисомы появляются при прорастании семян, в которых запасаются жиры, и содержат ферменты, необходимые для превращения жирных кислот в сахара: системы Р-окисления жирных кислот и глиоксилатный цикл (см. рис. 4.11). При работе ферментных систем пероксисом и глиоксисом образуется пероксид водорода, который разрушается содержащейся в этих органоидах каталазой.
Сферосомы. Это сферические, сильно преломляющие свет образования диаметром 0,5 мкм. Они содержат липиды и поэтому их называют также липидными каплями (олеосо-мами). В сферосомах обнаружены такие ферменты, как липаза и эстераза. В них хранятся запасы липидов клетки. При прорастании семян, запасающих жиры, сферосомы функционируют в комплексе с глиоксисомами в процессах глюконеогенеза.
Эндоплазматнческнй ретнкулум. Эндоплазматический ретику-лум (ЭР), или эндоплазматическая сеть (ЭС), представляет собой систему каналов, пузырьков и цистерн, ограниченную мембраной толщиной 5 — 6 нм. ЭР может содержать на своей поверхности рибосомы (гранулярный, или шероховатый, ЭР) или не содержать их (агранулярный, или гладкий, ЭР). Поверхность мембран ЭР в клетке больше других мембранных образований, а объем полостей может достигать 16% объема клетки. Эндоплазматическая сеть — очень лабильная структура. Неблагоприятные внешние воздействия (недостаток кислорода и др.) вызывают концентрические закручивания мембран ретикулума.
В мембранах ЭР локализованы редокс-цепи двух типов, с участием которых происходят детоксикация вредных для клетки соединений (NADPH-редуктаза и цитохром Р-450) и превращение насыщенных жирных кислот в ненасыщенные (NADH-редуктаза и цитохром Ь5). В гладком ЭР образуются углеводы, липиды, терпеноиды. В гранулярном ретикулуме синтезируются мембранные белки, ферменты, необходимые для синтеза полисахаридов клеточных стенок, Структурный белок и ферменты клеточных стенок, другие секретируемые белки. По системе ЭР переносятся вещества внутри клетки. Он участвует также в межклеточных взаимодействиях у растений через плазмодесмы (см. ниже).
Аппарат Гольджн. В растительных клетках аппарат Гольд-жи (АГ) представлен диктиосомами, везикулами и межцистер-ными образованиями. Уплощенные цистерны — диктиосомы расположены пачками по несколько штук. Они ограничены мембраной толщиной 7 — 8 нм. На регенерационном полюсе АГ происходит новообразование диктиосом из мембран гладкого ЭР. На секреторном полюсе формируются секреторные пузырьки (везикулы), содержащие предназначенные для секреишосшёсхва. В клетке растений содержатся от нескольких до сотен АГ.
В диктиосомах АГ образуются. гликодротеины. и гликоли-пиды и осуществляется накопление и мембранная «упаковка» соединений, необходимых для синтеза полимеров клеточной стенки и различных растительных слизей. С помощью везикул Гольджи углеводные компоненты доставляются к плазмалемме. Мембрана пузырьков встраивается в плазма-' лемму, способствуя ее росту и обновлению. Секретируемые вещества оказываются в клеточной стенке. Мембраны АГ являются связующим звеном между мембранами ЭР и плазмалеммой.
Вакуолярная система. Вакуоль — типичный органоид растительной клетки. В меристематических клетках вакуоли представлены мелкими пузырьками; для зрелых клеток характерна большая центральная вакуоль. Вакуолярная система растений формируется несколькими путями. Из расширенных цистерн ЭР образуются провакуоли, слияние которых приводит к возникновению более крупных вакуолей и созданию вакуо-лярной мембраны — тонопласта, который таким образом является производным ЭР. Тонопласт может образовывать инвагинации, что приводит к включению в вакуоль участков цитоплазмы. Гидролитические ферменты, содержащиеся в возникшей вакуоли, расщепляют полимеры до низкомолекулярных веществ.
Важную роль в становлении вакуЪлей играют явления автофагии. Процесс начинается с окружения мембраной ЭР участка цитоплазмы (автофагическая вакуоль). Деятельность кислых гидролаз в замкнутом мембраной пространстве приводит к деградации полимерного содержимого и поступлению воды. Вакуоли, возникшие в результате автофагии, могут сливаться с вакуолями, образовавшимися иными путями.
Вакуолярный сок имеет сложный состав и включает органические вещества и минеральные соли. Помимо органических кислот, углеводов, аминокислот и белков, которые могут быть вторично использованы в обмене веществ, клеточный сок содержит фенолы, таннины, алкалоиды, анто-цианы, которые выводятся из обмена веществ клетки в вакуоль и таким путем изолируются от цитоплазмы. Большинство ферментов вакуолей — гидролазы с оптимумом активности при кислом рН, что позволяет рассматривать вакуоли растительных клеток в качестве вторичных лизосом. Кислотность вакуолярного сока 5,0 — 6,5 единиц рН, но может быть равной 1,0 (бегония) или 2,0 (лимон).
Вещества доставляются в вакуоль с помощью разнообразных систем транспорта, функционирующих в тонопласте. К ним относится АТР-зависимая Н +-помпа, выносящая ионы Н + из цитоплазмы в вакуоль (см. рис. 1.3). Ее деятельность обеспечивает поступление в вакуоль анионов органических кислот, Сахаров, а также вход и выход ионов К + . Переносчики, локализованные в тонопласте, обусловливают накопление в вакуолях аминокислот и других соединений.
Важно отметить, что вакуоль может служить местом отложения запасных белков (алейроновые зерна). Процесс вакуолизации — необходимое условие роста клеток растяжением.
Мнкротрубочкн н мнкрофнламенты (цнтоскелет). В наружном кортикальном слое цитоплазмы неделящихся растительных клеток локализованы микротрубочки (см. рис. 13.2). Их наружный диаметр 30 нм, внутренний — около 14 нм. Они ориентированы параллельно друг другу и перпендикулярно продольной оси клетки (см. рис. 10.6). При делении клетки микротрубочки составляют основу структуры веретена, пучки трубочек прикрепляются также к кинетохорам хромосом. У монадных водорослей микротрубочки входят в состав жгутиков, обеспечивая их подвижность. Все микротрубочки имеют единый план строения и состоят из глобулярного кислого белка тубулина, субъединицей которого является димер, состоящий из двух глобулярных мономеров (а- и Р-тубулинов). В микротрубочке димеры белка располагаются по спирали. Трубка образована 13 субъединицами тубулина. Цитоплазматические микротрубочки легко диссоциируют на субъединицы (разбираются) и собираются вновь. Для сборки микротрубочек благоприятен кислый рН среды, присутствие магния, GTP и АТР. Разборка ускоряется повышением концентрации Са+ и низкой температурой.
В цитоплазме растительных клеток обнаружены также филаментные структуры, состоящие из немышечного актина. Это сократительный белок, сходный по молекулярной массе с актином мышц и близкий ему по аминокислотному составу. Он может находиться в мономерной (глобулярный, Г-актин) или в полимерной форме двойной спирали (фибриллярный, Ф-актин). Микрофиламенты актина взаимодействуют с микротрубочками кортикального слоя и плазмалеммой. Они участвуют в пространственной организации метаболических процессов, протекающих в растворимой фазе цитоплазмы, и служат основой ее двигательной активности.
Клеточная стенка. Клетки растений окружены плотной полисахаридной оболочкой, выстланной изнутри плазмалеммой. Формируется клеточная стенка на стадии телофазы во время митотического деления клеток. Клеточную стенку делящихся и растущих растяжением клеток называют первичной. После прекращения роста клетки на первичную клеточную стенку изнутри откладываются новые слои и возникает прочная вторичная клеточная стенка.
В состав клеточной стенки входят структурные компоненты (целлюлоза у растений, хитин у грибов), компоненты матрикса стенки (гемицеллюлозы, пектин, белки), инкрустирующие компоненты (лигнин, суберин) и вещества, откладывающиеся на поверхности стенки (кутин и воска). Клеточные стенки могут содержать также силикаты и карбонаты кальция.
Целлюлоза (полимер (З-Э-глюкозы), гемицеллюлозы (полимеры гексоз и пентоз) и пектиновые вещества (производные уроновых кислот) являются углеводными компонентами клеточных стенок. Целлюлоза и пектиновые вещества адсорбируют воду, обеспечивая оводненность клеточной стенки. Пектиновые вещества, содержащие много карбоксильных групп, связывают ионы двухвалентных металлов, которые способны обмениваться на другие катионы (Н + , К+ и т.д.). Это обусловливает катионообменную способность клеточных стенок растений. Помимо углеводных компонентов в состав матрикса клеточной стенки входит также структурный белок, называемый экстенсином. Это гликопротеин, содержащий более 20% L-оксийролина от суммы аминокислот. По этому признаку белок клеточных стенок растений сходен с межклеточным белком животных — коллагеном.
Основным инкрустирующим веществам клеточной стенки является лигнин. Интенсивная лигнифика^ия клеточных стенок начинается после прекращения роста клетки. Лигнин представляет собой полимер с неразветвленной молекулой, состоящей из ароматических спиртов (п-кумарового, кониферило-вого, синапового). Разрушение и конденсация лигнина в почве — один из факторов образования гумуса.
В регуляции водного и теплового режима растений участвуют ткани, стенки клеток которых пропитаны суберином. Отложение суберина делает стенки трудно проницаемыми для воды и растворов (например, в эндодерме, перидерме).
Поверхность эпидермальных клеток растений защищена гидрофобными веществами — кутином и восками: Предшественники этих соединений секретируются из цитоплазмы на поверхность, где и происходит их полимеризация. Слой кутина обычно пронизан полисахаридными компонентами стенки (целлюлозой, пектином) и образует кутикулу. Кутикула участвует в регуляции водного режима тканей и защищает клетки от повреждений и проникновения инфекции.
В первичных клеточных стенках на долю целлюлозы
Рис. 1.6
^
yj Схема
взаимосвязей между компонентами
первичной клеточной стенки (по W.
D.
Bauer,
1977)
Схема
взаимосвязей между компонентами
первичной клеточной стенки (по W.
D.
Bauer,
1977)
приходится до 30% сухой массы стенки. Количество геми-целлюлоз и пектиновых веществ меняется в зависимости от объекта. Вместе с белками пектиновые вещества могут составлять около 30% сухой массы клетки, причем количество белка достигает 5 — 10%. Около 40% приходится на долю гемицеллюлоз.
Каким образом взаимосвязаны рассмотренные компоненты в клеточной стенке? Молекулы целлюлозы взаимодействуют между собой водородными связями и собраны в микрофибриллы (рис. 1.6). Водородные связи существуют также между микрофибриллами целлюлозы и гемицеллюлозами. Большинство остальных связей в клеточной стенке ковалент-ные: между гемицеллюлозами и пектиновыми веществами, между пектиновыми веществами и экстенсином, между лигнином и целлюлозой, экстенсином и лигнином. В пектиновых полимерах, имеющих карбоксильные группы, существенную роль играют ионные связи с участием главным образом кальция. Эти взаимодействия обеспечивают прочность структуры клеточной стенки, ее эластичность и пластичность.
Благодаря контакту соседних клеток друг с другом возникает единая система клеточных стенок, получившая название апопласта. По апопласту, минуя мембранные барьеры, от клетки к клетке перемещаются вещества. Межмолекулярное пространство в фазе клеточной стенки, где осуществляются диффузия, адсорбция и освобождение водорастворимых веществ, называется кажущимся свободным пространством (см. 6.11.1).
Клеточные стенки растений пронизаны отверстиями — порами диаметром до 1 мкм. Через них проходят тяжи — плазмодесмы (рис. 1.7), благодаря которым осуществляются межклеточные контакты. Каждая плазмодесма представляет собой канал, выстланный плазмалеммой, непрерывно переходящей из клетки в клетку. Центральную часть поры занимает десмотрубка, состоящая из спирально расположенных белковых субъединиц. Десмотрубка сообщается с мембранами ЭР соседних клеток. Вокруг десмотрубки имеется слой цитоплазмы,
Рис. 17
 Схема
строения плаз-мод есмы :
Схема
строения плаз-мод есмы :
д_ продольный разрез; Б — поперечный разрез
1.2