

Сопряженная эволюция типов обмена веществ и среды обитания
вых количествах. В настоящее время он составляет 21 % по объему воздуха. Появление и накопление 02 в атмосфере связано с жизнедеятельностью зеленых растений. Ежегодно в ходе фотосинтеза кислород поступает в атмосферу в количестве 70—120 млрд. т. Этот кислород необходим для дыхания всех гетеротрофов — бактерий, грибов, животных и человека, а также зеленых растений в ночное время.
Особое значение в поддержании высокой концентрации 02 в атмосфере имеют леса. Подсчитано, что 1 га леса весной и летом за час выделяет 02 в количестве, достаточном для дыхания 200 человек.
5.Озоновый экран. Еще одно важнейшее следствие выделения растениями кислорода — образование озонового экрана в верхних слоях атмосферы на высоте около 25 км. Озон (03) образуется в результате фотодиссоциации молекул 02 под действием солнечной радиации. Озон задерживает большую часть ультрафиолетовых лучей (240 — 290 нм), губительно действующих на все живое. Возможность частичного разрушения озонового экрана из-за загрязнения атмосферы промышленными и другими отходами — серьезная проблема охраны биосферы.
Появление и эволюция живых организмов связаны с изменениями физико-химических условий на поверхности Земли. В свою очередь жизнедеятельность самих организмов оказывала и оказывает сильнейшее влияние на окружающую среду. Таким образом, система организмы — среда (т. е. биосфера) развивалась как единое целое. Последовательность этапов эволюции биосферы и их возраст можно представить в виде следующей гипотетической схемы (1 — условия среды, 2 — тип обмена веществ):
4,5
млрд. лет
4,0
млрд. лет
млрд. лет
Состав атмосферы: Н20, С02 и в небольших количествах N2, СН4, NH3, H2S и др. 02 практически отсутствует. Высокая температура, ультрафиолетовая радиация и электрические разряды способствуют синтезу органических веществ.
Абиотический синтез простых органических соединений и накопление их в первичном океане.
Снижение температуры, уменьшение облачности, появление следов 02 и озонового экрана из-за диссоциации паров воды под действием ультрафиолетовой радиации.
Образование сложных органических молекул: полипептидов, полисахаридов, нуклеиновых кислот, липидов, металлоорганических соединений с каталитическими и фотохимическими свойствами (в том числе металлопор-фиринов).
Образование предбиологических структур типа коа-церватов (А. И. Опарин) или протеиноидных микросфер (С. Фокс). Появление самовоспроизводящихся метаболических циклов (М. Эйген). Объединение всех этих компонентов в единой системе протобионтов.
1. Те же тенденции. Увеличение количества света, достигающего поверхности Земли.
2. Появление первичных анаэробных гетеротрофов, способных к брожению и обладающих окислительным пентозофосфатным циклом, которые питаются абиогенно синтетизированными органическими веществами. У некоторых из них свет с участием пигментов способствует окислению органических веществ (фотоорганотрофия); у других появляется способность в условиях гетеротрофного питания переносить электроны на сульфат или нитрат с участием цитохромов (хеморедукция).
3,0
млрд. лет
2,0-2,5 млрд. лет
Повышенное содержание С02, обусловленное жизнедеятельностью анаэробных гетеротрофов. Истощение абиогенно синтезированных органических веществ. Свет (400-800 нм). Следы 02.
Появление автотрофов, осуществляющих фоторедукцию (см. 3.3.1). В качестве легкоокисляемых веществ используются H2S, СН4, Н2 и др.
1. Те же тенденции. Истощение легко окисляющих- ся веществ, необходимых для фоторедукции.
2. Появление автотрофов-фотосинтетиков с системой нециклического фотофосфорилирования и механизмом фоторазложения воды (цианобактерии и зеленые водо- росли).
1,5
млрд. лет
1. Повышение содержания 02 и уменьшение кон- центрации С02 в атмосфере и гидросфере в результате деятельности фотосинтезирующих организмов.
2. Появление вторичных гетеротрофов (с аэробным типом дыхания) и аэробных хемосинтетиков.
Клеточный механизм фотосинтеза в ходе эволюции сформировался у одноклеточных организмов (у бактерий), причем цитохромы начали принимать участие в транспорте электронов, по-видимому, уже у первичных гетеротрофов. Как следует из приведенной схемы, сначала возник механизм циклического фотофосфорилирования (ФС I), а затем у цианобактерии — молекулярный комплекс нециклического фотофосфорилирования (ФС I + ФС II). Пентозофосфатный цикл окисления глюкозы также существовал у первичных гетеротрофов. Его обращение с использованием энергии света стало основным способом восстановления С02 у растений (цикл Кальвина).
Механизм фотосинтеза у растений сосредоточен в хлоропластах. Большой интерес представляет симбиотическая гипотеза происхождения хлоропластов, выдвинутая впервые А. С. Фаминцыным (1886, 1907) и поддержанная К. С. Мережковским (1905—1909). Согласно этой гипотезе, хлоропласты когда-то были самостоятельными одноклеточными фотосинте-зирующими организмами, которые жили в более крупной гетеротрофной клетке как симбионты. В процессе эволюции эти симбиотические отношения дали начало новой, более сложной клетке со специализированными и взаимодействующими органоидами. Фаминцын рассматривал процесс симбиогенеза как важнейший эволюционный путь усложнения клеток. Современные данные во многом подтверждают эту точку зрения. Хлоропласты имеют кольцевую ДНК и рибосомы прокариотического типа. Синтез белков в хлоропластах подавляется хлорамфениколом, как и v б ктепий. я не пт-пп-
гексимидом, который ингибирует процесс трансляции в цитоплазме эукариотических клеток. Хлоропласты способны к размножению путем простого деления.
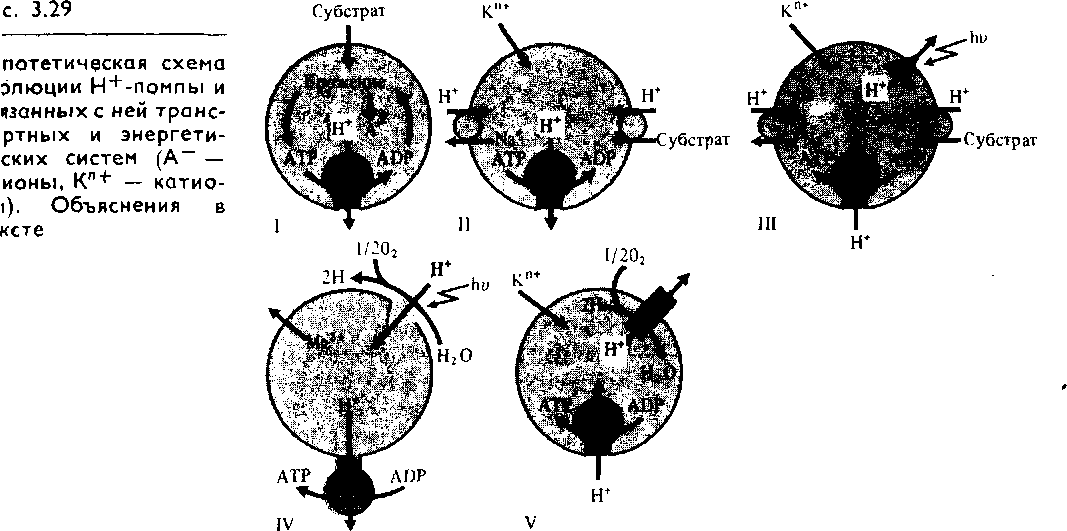
Решающее значение для совершенствования энергетических механизмов клеток имело появление в ходе эволюции способности к активному трансмембранному переносу ионов Н + . АТРазная Н*-помпа (рис. 3.29,1) должна была функционировать уже у самых примитивных клеток — протобион-тов — для удаления избытка ионов Н+, которые накапливались в них при брожении (анаэробном окислении глюкозы). В результате выкачивания ионов Н+ во внутриклеточной среде не только поддерживался оптимальный для синтетических процессов уровень рН, но и возникал электрохимический мембранный потенциал, который стал энергетической основой мембранного транспорта и осморегуляции (рис. 3.29, II).
Появление электрохимического мембранного потенциала ионов Н+, возможно, стало поворотным моментом в переходе протобионтов из неживого в живое состояние в соответствии с теорией советского биолога Э. С. Бауэра (1935), согласно которой живое состояние базируется на принципе устойчивого неравновесия.
При переходе бактериальных клеток на фототрофный способ питания роль Н+-помпы стала выполнять в клеточной мембране редокс-цепь. Примером такой светозависимой Н+-помпы является система бактериородопсина (производного каротина) у современных галофильных бактерий (рис. 3.29, III). Мембранный протонный градиент, возникший на мембране, способствовал обращению транспорта ионов Н+ через АТРазу и появлению фотофосфорилирования. Фотохимическая активность бактериородопсина не сопровождается диссоциацией воды и выделением 02.
Основой светозависимой Н+-помпы в растительном мире стала пигментная система, в состав которой входят хлоро
филлы. В результате образования тилакоидов путем инвагинации поверхностной мембраны у предшественников хлоропластов (симбионтов) Н+-помпа переносит ионы Н+ во внутреннее пространство тилакоидов (рис. 3.29, IV). Пассивный выход ионов Н+ через протонный канал АТРазы по концентрационному градиенту сопровождается синтезом АТР (см. рис. 4.8).
В клеточной мембране аэробных гетеротрофных бактерий и во внутренней мембране митохондрий1 эукариотических клеток в качестве Н+-помпы функционирует дыхательная электронтранспортная цепь, выкачивающая ионы Н+ наружу (рис. 3.29, V). В результате обратного пассивного транспорта протонов синтезируется АТР (окислительное фосфори-лирование). Таким образом, приспособительная эволюция энергетических систем живых организмов, а вместе с тем и газообмена клеток с окружающей средой связана с преобразованием механизмов Н+-помпы.
В ходе дальнейшей эволюции растений (появление многоклеточное™, выход на сушу) внутриклеточная организация процессов фотосинтеза изменялась незначительно. Примером таких изменений может служить появление С4-пути фотосинтеза и САМ-метаболизма. Прогрессивное развитие функции фото-трофного питания у многоклеточных растений в основном происходило за счет анатомо-морфологических приспособлений. Главное из этих приспособлений — появление листа как специализированного органа фотосинтеза и совершенствование его анатомической структуры.
В ходе световой фазы фотосинтеза в мембранах тилакоидов хлоропластов энергия света используется для окисления Н2О (с выделением 02), восстановления NADP+ и образования трансмембранного электрохимического потенциала ионов Н + , который трансформируется в энергию высокоэнергетической фосфатной связи АТР (фото-фосфорилирование). Молекулы хлорофилла, поглощающие красный и синий свет, выполняют функцию фотосенсибилизаторов. Каротиноиды защищают хлорофилл от разрушения, а также, как и фико-билины у водорослей, служат дополнительными пигментами. В основе световой фазы фотосинтеза лежит механизм Н+-помпы.
В ходе темновой фазы фотосинтеза энергия АТР и NADPH расходуется на восстановительное карбоксилирование (ассимиляцию С02) с образованием 3-фосфоглицеринового альдегида (Сз-путь фотосинтеза) или органических кислот (С4-путь) в качестве первичных продуктов. Энергия света в хлоропластах может использоваться также на восстановление NOj" и S04~. При недостатке С02 и избытке 02 включается фотодыхание с участием пероксисом.
1
Предполагается, что митохондрии могли
произойти от аэробных бактерий
также путем симбиогенеза.
Г" л а в а 4
ДЫХАНИЕ РАСТЕНИЙ
«...Дыхание служит стимулом главнейших жизненных отправлений растений ... подобно животным, они поглощают при переработке органических соединений в органические образования кислород, выделяют углекислоту и освобождают тепловую энергию...»
А. С. Фаминцын
4.1