

9.6. Интраорганная нервная система и тканевые рецепторы
А. Общая характеристика
1. Внутренние органы после экстраорганной денервации — перерезки симпатических и парасимпатических нервов, продолжают функционировать. Вначале наблюдаются незначительные дистрофические явления, которые скоро исчезают. Через несколько недель чувствительность внутренних органов к биологически активным веществам и в первую очередь к медиаторам повышается, их функция улучшается. Многие внутренние органы продолжают функционировать, будучи изолированными, находясь в физиологическом растворе: сокращаются перфузируемое сердце, мочеточники, желчный пузырь, кишечник. В пищеварительном тракте после перерезки симпатических и парасимпатических путей сохраняются все виды двигательной активности: перистальтика, ритмическая сегментация, маятникообразные движения, продолжается всасывание. Двигательная активность пищеварительных органов после перерезки их нервов регулируется рефлекторными дугами, замыкающимися в пределах мышечного и подслизистого сплетений его стенок, а также в экстраорганных ганглиях. Возбуждение афферентных нейронов кишечной стенки под действием пищевого комка приводит к возникновению рефлексов, при которых пищевой комок продвигается в кау-дальном направлении путем сокращения мышц орального участка кишки и одновременного расслабления каудального участка. Дуги этих двух рефлексов, обусловливающих перистальтику, лежат в пределах стенки кишечника. Тормозные нейроны этих дуг не являются ни холинергическими, ни адренер-гическими; возможно, их медиатором служит АТФ. Возбуждающие нейроны выделяют аце-тилхолин, хотя не исключено, что их действие может быть опосредовано и другими медиаторами, например серотонином. Имеются спонтанно активные эфферентные нейроны; на них могут конвергировать интра- и экстраорганные нервные волокна.
167
-
Интраорганная нервная система содер жит все элементы рефлекторной дуги: аффе рентный, вставочный и эффекторный нейро ны. Чувствительные интрамуральные нейро ны — клетки Догеля II типа; они образуют первое звено — рецептор, и второе — аффе рентный путь рефлекторной дуги. Показано наличие механо-, хемо- и термочувствитель ных клеток. Обнаружены быстро- и медлен но адаптирующиеся тонические нейроны, возбуждающие и тормозящие сокращения кишки.
-
На одну и ту же клетку интрамураль- ных ганглиев конвергируют межганглионар' ные (вставочные) и экстраорганные (по- стганглионарные симпатические и прегангли- опорные парасимпатические волокна). Пара симпатические преганглионарные волокна образуют синапсы на возбуждающих и, по- видимому, тормозящих нейронах. Симпати ческие постганглионарные волокна оказыва ют непосредственное тормозное влияние на гладкую мышцу кишки, а также тормозят хо- линергические ганглионарные нейроны, уг нетают выделение медиатора из пресинапти- ческих окончаний преганглионарных пара симпатических волокон. Местные рефлек торные дуги выявлены и в сердце. Плотность расположения интраорганных нейронов весьма высока — в кишечнике, например, находится около 20 тыс. нейронов на 1 см2. В целом в кишке число нейронов (МО8) пре восходит таковое в спинном мозге.
Б. В интраорганной нервной системе богат набор медиаторов и соответствующих рецепторов. Только в энтеральной ее части функционирует около 20 различных медиаторов и модуляторов. Наиболее изучены ацетилхо-лин, катехоламины, серотонин, ГАМК, пептиды, АТФ. В частности, серотонин обнаружен в различных тканях организма животных. Однако больше всего серотонина (90 % от его общего количества, содержащегося в организме) приходится на энтерохромаф-финные клетки пищеварительного тракта. Серотонин вызывает сокращение гладких мышц внутренних органов, в том числе кровеносных сосудов и бронхов. При прямом действии серотонина возрастает сила сердечных сокращений, хотя этот эффект маскируется рефлекторными барорецептивными влияниями противоположного направления. В центральном и периферическом отделах нервной системы выявлено несколько типов серотонинорецепторов: 5-НТ,—5-НТ5 (S,— S5 — по первой букве названия медиатора по аналогии с названием гистаминорецепто ров - Н, и Н2, дофаминорецепторов — D,
пуринорецепторов — Р, и Р2, N- и М-холино-рецепторов). 5,,2-рецепторы находятся в гладкой мышце желудочно-кишечного тракта, 53-рецепторы — в вегетативных ганглиях, S4,5-peuenTopbi обнаружены в ЦНС. Установлено наличие не только пост-, но и преси-наптических серотонинорецепторов.
Медиаторная роль АТФ и ее производных в настоящее время активно изучается. Опыты обычно ставят на изолированной мышечной полоске кишки. Считают, что в интрамураль-ном нервном аппарате имеются пуринергические нейроны, аксоны которых выделяют пуриновые продукты (отсюда и название нейронов) распада АТФ — инозин и адено-зин. АТФ оказывает тормозящее влияние на моторику желудочно-кишечного тракта. Активация пуринергических структур вызывает расслабление кишки, а возникающие вслед за этим ее сокращения под влиянием холи-нергических нейронов обеспечивают прохождение болюса по кишке (пропульсия). Пуринергические нейроны участвуют также в механизме релаксации желудка, расслаблении пищеводного и анального сфинктеров. Выделено два типа пуринорецепторов (Р, и Ра); рецепторы первого типа более чувствительны к аденозину, второго — к АТФ. Р,-ре-цепторы преобладают в сердечно-сосудистого системе, трахее, мозге. Они связаны со специфической аденилатциклазой. Р2-рецепто-ры расположены в основном на клетках органов желудочно-кишечного тракта и мочеполовой системы. Тормозящее (гиперполяри-зующее) влияние АТФ реализуется, по-видимому, через повышение проницаемости клеточных мембран для ионов калия. Однако для желудочно-кишечного тракта медиаторная роль АТФ убедительно не доказана.
Гистаминорецепторы (подтипы Н, и Нг) имеются на клетках всех органов и участвуют в регуляции различных функций организма (табл. 9.2).
Дофаминореиепторы: тип D, выявлен в симпатических ганглиях, паращитовидных железах, по-видимому, все дофаминрецепто-ры являются постсинаптическими. Drpe-цепторы найдены на мембранах лимфоцитов. D,- и D2-peuenTopbi более широко распространены в ЦНС.
ВИП-рецепторы (ВИП — вазоактивный интестинальный пептид) вызывают накопление цАМФ и усиление моторной, секреторной активности, кровоснабжения желудка и кишечника.
Рецепторы вещества П (ВП): имеются на гладкомышечных клетках кишки, клетках мочевого пузыря, семявыносящих протоков.
168
Таблица 9.2. Гистамияовые рецепторы, эффекты их активации
|
Тип гис-таминовых рецепторов |
Локализация рецепторов |
Эффекты |
|
Hi |
Гладкие мышцы бронхов Гладкие мышцы кишечника Сердце Гладкие мышцы артериальных сосудов Гладкие мышцы вен Капилляры |
Повышение тонуса Повышение тонуса Угнетение атрио-вентрикулярной проводимости Снижение тонуса Повышение тонуса Повышение проницаемости |
|
н2 |
Париетальные клетки слизистой оболочки желудка Сердце Глаакие мышцы артериальных сосудов Миометрий |
Повышение секреторной активности Увеличение частоты и силы сокращений Снижение тонуса Угнетение сократительной активности |
Далеко не все рассмотренные рецепторы иннервируются интра- или экстраорганной нервной системой; иннервацию имеют М-хо-лино-, а,- и р1,-, серотонино-, пуринорецеп-торы. Степень иннервации других рецепторов изучена мало. Активность окончаний экстра- и интраорганной нервной системы регулируется количеством выделившегося из них медиатора, а также другими биологически активными веществами.
9.7. РЕГУЛЯЦИЯ ФУНКЦИЙ СИНАПСОВ
Эффективность передачи в синапсе ВНС зависит от количества медиатора, выделяемого в синаптическую щель, которое, во-первых, определяется числом импульсов, пришедших ктерминали, — с увеличением импульсации выделение медиатора до определенного значения увеличивается, при ослаблении импульсации выделение медиатора уменьшается. Во-вторых, важную роль в регуляции выделения медиаторов в синаптическую щель постганглионарными окончаниями ВНС играют пресинаптические адрено- и холиноре-
цепторы, имеющиеся на самих окончаниях. Эти рецепторы реализуют свое влияние по принципу отрицательной обратной связи, вызывающей противоположные эффекты: малые порции медиатора стимулируют его выделение, большие — угнетают.
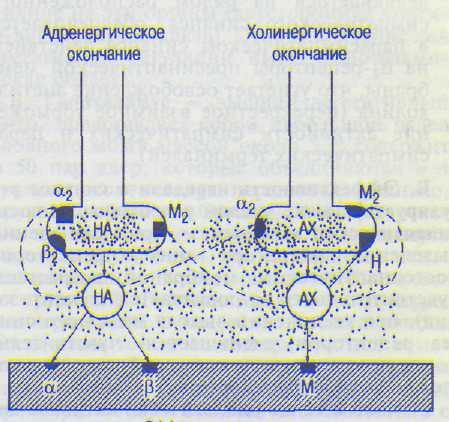
А. Регуляция выделения нор адреналин а. На симпатических постганглионарных окончаниях имеются а2- и pV^PeH0PeUenT°Pbi и М2-холинорецепторы. Активация М2-холи-норецепторов и сс2-адренорецепторов тормозит, а р2-адренорецепторов стимулирует освобождение медиатора (рис. 9.7).
Пресинаптические руадренорецепторы более чувствительны: они активируются низкими концентрациями норадреналина и усиливают его секрецию. Пресинаптические а2-адренорецепторы менее чувствительны: они активируются высокими концентрациями норадреналина и уменьшают его секрецию нервными окончаниями. Эффекторные клетки с помощью образуемых ими проста-гландинов группы Е также тормозят освобождение норадреналина из пресинаптичес-ких окончаний. Активация пресинаптичес-ких М2-холинорецепторов осуществляется ацетилхолином, выделяющимся из рядом расположенных парасимпатических оконча-
Эффекторная клетка
Рис. 9.7. Саморегуляция секреции медиатора симпатическими и парасимпатическими постганглионарными нервными окончаниями с помощью пре-синаптических аз- и fb-адренорецепторов, М2- и Н-холинорецепторов и их реципрокные взаимодействия.
Сплошные линии — усиление, пунктирные —- угнетение выделения медиатора- НА — норадреналин; АХ — ацетил-холин.
169
ний. На окончаниях преганглиоиарных симпатических волокон имеются рецепторы к се-ротонину, гистамину, кортикостероидам и Н-холинорецепторы, активация которых облегчает синаптическую передачу. М|-холино-рецепторы, рецепторы дофамина, наоборот, обеспечивают ингибирование секреции аце-тилхолина преганглионарными симпатическими окончаниями.
Б. Регуляция освобождения ацетилхолина из пресинаптических окончаний лостгапглио-нарных волокон осуществляется также несколькими механизмами.
-
По принципу обратной связи — с помо щью взаимодействия выделившегося в си наптическую щель ацетилхолина с М2- и Н-холинорецепторами пресинаптической мембраны: связывание с М2-холинорецеп> торами тормозит, связывание с Н-холино рецепторами усиливает выделение медиа тора.
-
Ретроингибирование с помощью АТФ обеспечивается пуринорецепторами пре синаптической терминали. Большое коли чество АТФ выделяется в синаптическую щель под влиянием ацетилхолина из эф- фекторной клетки.
-
Тормозящее влияние норадреналина, вы деляющегося из рядом расположенного симпатического синапса, обеспечивается в парасимпатическом синапсе действием на а.2-рецепторы пресинаптической мем браны, что угнетает освобождение ацетил холина (перекрестное взаимное торможе ние активности симпатических и пара симпатических терминален).
В. Эффективность передачи в синапсе регулируется также числом рецепторов на пост-синаптической мембране — при уменьшении
выделения медиатора синтез рецепторов постсинаптической мембраной возрастает, чувствительность ее повышается (сенситиза-ция), при увеличении выхода медиатора синтез рецепторов уменьшается, чувствительность постсинаптической мембраны снижается — десенситизация.
Г. Одно и то же биологически активное вещество может выполнять функции медиатора и модулятора. Например, ацетилхолин и но-радреналин, выделяемые из постганглионар-ных нервных окончаний, не только действуют на рецепторы эффекторных клеток, но одновременно регулируют выделение медиатора самими нервными окончаниями — модулирующая роль. Некоторые вещества, по-видимому, играют роль только модуляторов функции нервных окончаний и эффекторных
клеток. Наиболее важными нейромодулято-рами являются кортикостероиды, половые гормоны, АКТГ, соматостатин, ангиотензин, эндорфин, энкефалины, простагландины, Взаимодействие нейромодуляторов с клеточными внесинаптическими рецепторами вызывает изменение либо проницаемости клеточной мембраны, либо клеточного метаболизма в результате их проникновения в цитоплазму клеток. В обоих случаях это ведет к усилению влияния нервных волокон на эф-фекторную клетку.