

2. Установлена определенная зависимость между содержанием йода и гормонообразова-
187
тельной активностью щитовидной железы.
Малые дозы йода стимулируют образование гормонов за счет усиления функции секреции фолликулов железы. Большие дозы йода по принципу обратной отрицательной связи тормозят процессы гормонопоэза, особенно когда функция железы повышена.
-
Вегетативная нервная система: воз буждение постганглионарных симпатических нервных волокон, идущих от средних шей ных симпатических ганглиев, приводит к по вышению активности щитовидной железы, а активность блуждающего нерва обусловлива ет снижение гормонообразовательной функ ции железы.
-
Гипоталамическая область также ока зывает выраженное влияние на образование гормонов в щитовидной железе через тирео- либерин (см. рис. 3.1).
-
Возбуждение ретикулярной формации ствола мозга ведет к повышению функцио нальной активности щитовидной железы за счет усиления выработки тиреолиберина ги поталамусом.
-
Кора большого мозга также принимает участие в регуляции активности щитовидной железы. В опытах на животных с удаленной корой было установлено, что в первый пери од после декортикации отмечается усиление активности щитовидной железы, в дальней шем функция железы снижается.
Б. Транспорт. Оба йодированных гормона в крови находятся не в свободном виде, а в соединении с белками глобулиновой фракции. Этот белок получил название тироксин-связывающий глобулин. Кроме того, гормоны щитовидной железы могут связываться также альбумином плазмы крови. Тироксин прочнее комплексируется с белками плазмы крови, чем трийодтиронин. Поэтому последний быстрее и лучше проникает в ткани и проявляет в них большую биологическую активность, чем тироксин. Для исследования в крови циркулирующих гормонов щитовидной железы определяют связанный с белком йод (СБЙ), так как йод входит в основном в состав тиреоидных гормонов. В норме уровень СБЙ составляет 4—8 мкг%. При поступлении тироксина в кровоток он захватывается, в частности, клетками печени. Печень является главным органом, регулирующим уровень гормонов щитовидной железы. В печени тироксин образует парные соединения с глю-куроновой кислотой. Последние не обладают гормональной активностью и выносятся желчью в желудочно-кишечный тракт, а далее удаляются. Образование парных соединений тироксина с глюкуроновой кислотой рас-
сматривается как дезинтоксикационный процесс, благодаря которому предотвращается чрезмерное насыщение крови гормонами. Опыты с радиоактивным 1 показали, чтоб организме взрослого человека ежесуточно полностью разрушается в среднем около 300 мкг тироксина и трийодтиронина.
10.6.2. ФИЗИОЛОГИЧЕСКАЯ РОЛЬ ЙОДИРОВАННЫХ ГОРМОНОВ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
Иодированные гормоны щитовидной железы оказывают выраженное влияние на многие функции и ткани организма.
-
Влияние на функции ЦНС было, в част ности, продемонстрировано в опытах на со баках при длительном введении больших доз тироксина — животные становятся беспо койными, у них усиливаются сухожильные рефлексы, появляется дрожание конечнос тей. Удаление щитовидной железы у живот ных резко снижает их двигательную актив ность, ослабляет оборонительные реакции. Введение тироксина устраняет указанные на рушения.
-
Влияние на высшую нервную деятель ность, У собак после удаления щитовидной железы условные рефлексы и дифференциро- вочное торможение вырабатываются с боль шим трудом. Сформированный условный рефлекс оказывается на следующий день ут раченным, и его приходится вырабатывать снова. Введение тироксина усиливает про цесс возбуждения в коре большого мозга, что приводит к нормализации условнорефлек- торной деятельности животных.
-
Влияние на процессы роста и развития показано в различных опытах и клинических наблюдениях. Так, удаление щитовидной же лезы в молодом возрасте вызывает задержку роста тела. Нарушается развитие скелета. Центры окостенения появляются поздно. За медляется развитие почти всех органов, по ловых желез.
-
Влияние на обмен веществ. Тиреоидные гормоны воздействуют на обмен белков, жиров, углеводов, минеральный обмен. Ти роксин усиливает расходование всех видов питательных веществ, повышает потребление тканями глюкозы. Под влиянием гормонов щитовидной железы заметно уменьшаются запасы жира и гликогена в печени. Многооб разное действие йодированных гормонов на обмен веществ связано с их влиянием на внутриклеточные процессы окисления и об разования белка. Усиление энергетических
188

5. Влияние на деятельность органов. Тироксин увеличивает частоту сердечных сокращений, дыхательных движений, повышает потоотделение. Кроме того, тироксин снижает способность крови к свертыванию и повышает ее фибринолитическую активность. Это связано с тем, что гормон уменьшает образование в печени, почках, легких и сердце факторов, принимающих участие в процессе свертывания крови, и увеличивает синтез антикоагулянтов, а также веществ, повышающих фибринолитические свойства крови. Тироксин угнетает функциональные свойства тромбоцитов — их адгезивную и агрегаци-онную способность, что обусловлено изменением в них метаболизма простагландинов и циклических нуклеотидов.
При недостаточности функции щитовидной железы (гипотиреоз) в детском возрасте возникает кретинизм. При недостаточной функции щитовидной железы может возникнуть и другое патологическое состояние, которое получило название «микседема» (термин греческого происхождения — слизистый отек). Заболевание редкое, встречается преимущественно в детском возрасте, у стариков, а также у женщин в климактерическом периоде. При повышении функциональной активности щитовидной железы (гипертире-оз) возникает патология — тиреотоксикоз (базедова болезнь). В некоторых местностях в воде отмечается недостаток йода. Это приводит к снижению функции щитовидной железы со значительным компенсаторным разрастанием ее ткани, образующим так называемый зоб. Заболевание, возникающее при этом, получило название эндемического зоба.
10.6.3. ФИЗИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ ТИРЕОКАЛЬЦИТОНИНА
Гормон образуется парафолликулярными клетками щитовидной железы, которые расположены вне ее железистых фолликулов. Под влиянием тиреокальцитонина снижается уровень кальция и фосфатов в крови (схема 10.1).
Понижение концентрации кальция в крови под влиянием кальцитонина происхо-
дит потому, что он тормозит выведение ионов кальция из костной ткани и увеличивает его отложение в ней. Тиреокальцитонин угнетает функцию остеокластов, разрушающих костную ткань, и активирует функцию остеобластов, принимающих участие в образовании новой костной ткани. Уменьшение содержания ионов кальция и фосфата в крови после введения тиреокальцитонина обусловлено снижением канальцевой реаб-сорбции ионов кальция и фосфата в почке, что увеличивает выделение их с мочой. Тиреокальцитонин активирует кальциевый насос клеточной мембраны, что способствует выходу ионов кальция из клетки. Кроме того, гормон прямым или косвенным образом стимулирует поглощение ионов кальция органел-лами клеток. За счет этих двух процессов происходит снижение концентрации ионов кальция в цитоплазме клеток.
Ключевую роль в регуляции секреции тиреокальцитонина играет уровень ионов циркулирующего кальция: повышение его концентрации быстро приводит к дегрануляции парафолликулярных клеток, появлению в них экзоцитоза, что сопровождается повышением содержания тиреокальцитонина в крови (см. схему \0Л). Активная секреция
189
тиреокальцитонина в ответ на гиперкальцие-мию способствует поддержанию концентрации ионов кальция в крови на определенном уровне (2,25—2,75 ммоль/л, или 9—11 мг%). Способствуют секреции тиреокальцитонина некоторые биологически активные вещества — гастрин, глюкагон, холецистокинин. Активация бета-адренорецегтторов повышает, а альфа-адренорецепторов — угнетает секрецию тиреокальцитонина.
10.7. ПАРАЩИТОВИДНЫЕ ЖЕЛЕЗЫ
Паращитовидные железы расположены на поверхности или погружен ы внутрь щитовидной железы. Паращитовидные железы хорошо снабжаются кровью. Они имеют как симпатическую (от шейных ганглиев), так и парасимпатическую (блуждающий нерв) иннервацию. Паращитовидные железы вырабатывают гормон, который получил название паратгормон (паратирин). Паратгормон выделяется в кровь в виде прогормона, превращение которого в гормон происходит в комплексе Гольджи клеток органов-мишеней в течение 15—30 мин.
Паратгормон наряду с кальцитонином регулирует обмен кальция в организме и поддерживает постоянство его уровня в крови.
При усилении деятельности паращитовид-ных желез (гиперпаратиреоз) наблюдается повышение концентрации кальция в крови. Наоборот, при недостаточности паращито-видных желез (гипопаратиреоз) происходит значительное снижение уровня кальция в крови. Известно, что костная ткань скелета является главным депо кальция в организме, поэтому имеется определенная зависимость между уровнем кальция в крови и содержанием его в костной ткани. Под влиянием паратгормона наблюдается усиление активности остеокластов, что приводит к высвобождению ионов кальция и фосфата из минерального вещества, образующего кость (гидроксиапатита) и поступлению их в кровь (схема 10.2). Паратгормон также активирует связанный с мембраной костных клеток фермент аденилатциклазу и за счет этого увеличивает поступление ионов кальция в кровоток. Кроме того, он усиливает реаб-сорбцию ионов кальция в почке, способствуя, таким образом, повышению уровня кальция в крови. Влияя на обмен кальция, паратгормон одновременно воздействует на обмен фосфора в организме: уменьшает обратное всасывание фосфатов в дистальных канальцах почек, что приводит к большему выделению с мочой и понижению их концентрации в крови.
Схема 10.2. Физиологическая роль и регуляция секреции паратирина
р-Адрено-рецепторы
Аденилатциклзза
а-Адрено-рецепторы
ц-АМФ
X
Паращитовидные железы
i.
Паратирин

остеокластов
Снижение активности
остеокластов
Ворсинки
слизистой
ТОНКОЙ КИШКИ
![]() Усиление
выхода
Са2+
Усиление
выхода
Са2+
в кровь из костной ткани
Уменьшение поступления Са2+
из крови в костную ткань
Усиление реабсорбции Са2+
Усиление всасывания Са2+в кровь
Гиперкальциемия (более 2,75 ммоль/л}
Гипокальциемия (менее 2,25 ммоль/л)
190
Паратгормон увеличивает всасывание ионов кальция и неорганического фосфата из кишечника, что обусловлено усилением поглощения клетками ворсинок слизистой кишечника этих ионов и последующим переходом их в кровь. После удаления паращито-видных желез в крови снижается уровень кальция и возрастает содержание фосфатов. Следовательно, между концентрацией кальция и фосфатов в крови существуют обратные соотношения.
Необходимо отметить, что гипофиз не вырабатывает гормонов, которые могли бы повлиять на функции паращитовидных желез. Регуляция активности этих желез определяется уровнем ионов кальция в крови. Если в крови концентрация ионов кальция нарастает, то это приводит к снижению функциональной активности паращитовидных желез. При уменьшении же уровня кальция в крови происходит повышение гормонообразова-тельной функции паращитовидных желез — принцип обратной отрицательной связи. Регуляция секреции паратгормона концентрацией ионов кальция в сыворотке крови основана на взаимодействии этого иона как с кальциевым рецептором кальмодулином, так и с альфа- или бета-адренорецепторами мембраны клеток паращитовидной железы. В результате этого взаимодействия изменяются активность аденилатциклазы и образование циклического аденозинмонофосфата (цАМФ). Это приводит к немедленному изменению скорости секреции паратгормона.
Удаление паращитовидных желез у животных или их недостаточная функция у человека приводят к развитию вялости, потере аппетита, рвоте, разрозненным сокращениям отдельных мышц (фибриллярные подергивания), в тяжелых случаях переходящим в длительные сокращения (тетанию — механизм см. 4.2.2). Этот процесс преимущественно захватывает мышцы конечностей, лица и затылка. Характерным симптомом у человека является так называемая «рука акушера», так как тонус мышц сгибателей кисти сильнее, чем тонус разгибателей. В тяжелых случаях спазм гортани, паралич дыхательных мышц и остановка сердца приводят к смерти.
10.8. ВИЛОЧКОВАЯ ЖЕЛЕЗА (ТИМУС)
Вилочковая железа — парный дольчатый орган, расположенный в верхнем отделе переднего средостения. Она состоит из двух долей неодинаковой величины, соединенных между собой прослойкой соединительной
ткани. Иннервация тимуса осуществляется парасимпатическими (блуждающими) и симпатическими нервами, берущими начало от нижнего шейного и верхнего грудного симпатического ганглиев. Вилочковая железа образует несколько гормонов: тимозин, гомео-статический тимусный гормон, тимопоэтин I, тимопоэтин II и тимусный гуморальный фактор. Все они являются полипептидами. Гормоны вилочковой железы играют большую роль в развитии иммунологических защитных реакций организма, стимулируя образование антител, которые обеспечивают реакцию организма на чужеродный белок.
Тимус контролирует развитие и распределение лимфоцитов, участвующих в иммунологических реакциях. Эта функция осуществляется либо путем насыщения лимфоидной ткани лимфоцитами, либо за счет выработки гормонов, стимулирующих развитие этих клеток крови, необходимых для обеспечения защитных иммунологических реакций. Недифференцированные стволовые клетки, которые образуются в костном мозге, выходят в кровоток и поступают в вилочковую железу. В тимусе они размножаются и дифференцируются в лимфоциты тимусного происхождения (Т-лимфоциты). Полагают, что именно эти лимфоциты ответственны за развитие клеточного иммунитета. Т-лимфоциты составляют большую часть циркулирующих в крови лимфоцитов (60—80 %).
Секреция гормона тимуса регулируется системой гипоталамус — передняя доля гипофиза. Соматотропин (гормон роста) способствует поступлению гормонов вилочковой железы в кровь.
Вилочковая железа достигает максимального развития в детском возрасте. После наступления полового созревания она останавливается в развитии и начинает атрофироваться. В связи с этим полагают, что железа стимулирует рост организма и тормозит развитие половой системы.
Физиологическое значение вилочковой железы связано с тем, что она содержит в своих клеточных структурах большое количество витамина С, уступая в этой отношении только надпочечникам.
При увеличении вилочковой железы у детей возникает тимико-лимфатический статус. Считают, что указанное состояние является врожденной конституциональной особенностью организма. При этом статусе, кроме увеличения тимуса, происходит разрастание лимфатической ткани. Характерен внешний вид больного: пастозное, одутловатое лицо, рыхлость подкожной клетчатки,
191
тучность, тонкая кожа, мягкие волосы. Существует мнение о том, что увеличение ви-лочковой железы у детей — проявление над-почечниковой недостаточности.
Клинически тимико-лимфатический статус может проявляться в повторной беспричинной рвоте, изменении дыхания и даже сердечно-сосудистом коллапсе (острая сердечная недостаточность). В тяжелых случаях может наступить смерть.
10.9. ПОДЖЕЛУДОЧНАЯ ЖЕЛЕЗА
10.9.1. СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ХАРАКТЕРИСТИКА
Поджелудочная железа относится к железам со смешанной функцией. Ацинозная ткань этой железы вырабатывает пищеварительный поджелудочный сок, который через выводной проток выделяется в полость двенадцатиперстной кишки. Внутрисекреторная деятельность поджелудочной железы проявляется в ее способности образовывать гормоны, которые поступают из железы непосредственно в кровь.
Впервые на эндокринную роль поджелудочной железы обратили внимание немецкие ученые Дж.Меринг и О.Минковский (1889). Они обнаружили, что после удаления у собак поджелудочной железы развиваются симптомы, которые отмечаются у человека при заболевании сахарным диабетом: резко увеличивается уровень сахара в крови, он выделяется в значительных количествах с мочой; появляются повышенный аппетит, жажда, происходит усиленное отделение мочи. Если таким животным пересадить под кожу поджелудочную железу, то все отмеченные изменения исчезают. В 1901 г. русский врач Л.В.Соболев подтвердил данные о том, что поджелудочная железа выполняет эндокринную функцию. По мнению Л.В.Соболева, поджелудочная железа выделяет гормоны, которые принимают участие в регуляции углеводного обмена. Однако в течение длительного времени эти гормоны не могли выделить из ткани поджелудочной железы. Это связано с тем, что гормоны поджелудочной железы являются полипептидами, которые разрушаются под влиянием протеолитических ферментов поджелудочного сока. Л.В.Соболевым впервые были предложены два метода получения гормона поджелудочной железы — инсулина. При первом способе у животного за несколько дней до удаления поджелудочной железы перевязывают выводной проток. При этом не происходит выделения поджелудочного сока в полость двенадцатиперстной кишки, ацинозная ткань атрофируется. В результате исключается возможность воздействия ферментов поджелудочного сока на инсулин и он может быть выделен из ткани железы. Кроме того,
Л.В.Соболев предложил извлекать инсулин из поджелудочной железы эмбрионов и новорожденных телят. В этот период поджелудочная железа еше не образует пищеварительного сока, но синтез инсулина уже осуществляется. Спустя 20 лет после публикации этих работ канадские ученые Ф.Бантинг и С.Бест получили активные препараты инсулина.
Морфологическим субстратом эндокринной функции поджелудочной железы служит островковый аппарат поджелудочной железы (островки Лангерганса), разбросанные среди ацинозной ткани железы. Островки расположены неравномерно по всей железе. Они преимущественно находятся в ее хвостовой части, и только небольшое количество их имеется в головном отделе железы.
У человека на 1 г железы приходится 3-25 тыс. островков Лангерганса. Островки Лангерганса состоят из альфа- (А), бета- (В), дельта-, РР- и G-клеток. Основную массу островков Лангерганса составляют бета-клетки. Около Vs общего количества клеток приходится на долю альфа-клеток. Последние по своим размерам крупнее бета-клеток и расположены преимущественно по периферии островка.
В бета-клетках образуется инсулин из своего предшественника — проинсулина. Синтез последнего осуществляется в эндоплазмати-ческом ретикулуме островковых клеток. Затем он переносится в клеточный аппарат Гольджи, где происходят начальные стадии превращения проинсулина в инсулин. Альфа-клетки синтезируют глюкагон, дельта-клетки — соматостатин. РР-клетки образуют в небольшом количестве панкреатический полипептид — антагонист холецистокинина. G-клетки вырабатывают гастрин. В эпителии мелких выводных протоков происходит образование липокаической субстанции, которую одни исследователи относят к панкреатическим гормонам, другие рассматривают ее как вещество энзиматической природы.
Поджелудочная железа иннервируется симпатическими и парасимпатическими нервами. Симпатические нервы представлены волокнами, идущими из солнечного сплетения, парасимпатические — блуждающим нервом. Их роль заключается в регуляции как образования и секреции гормонов, так и кровоснабжения поджелудочной железы.
Гистохимически установлено, что в ост-ровковой ткани железы содержится большое количество цинка. Цинк является и составной частью инсулина. Поджелудочная железа имеет обильное кровоснабжение.
192
10.9.2. ГОРМОНЫ ПОДЖЕЛУДОЧНОЙ ЖЕЛЕЗЫ
Инсулин принимает участие в регуляции углеводного обмена. Под действием этого гормона происходит уменьшение концентрации глюкозы в крови (в норме содержание глюкозы в крови 4,45—4,65 ммоль/л, или 80— 120 мг%) — возникает гипогликемия. Понижение уровня глюкозы в крови под влиянием инсулина связано с тем, что гормон способствует превращению глюкозы в гликоген в печени и мышцах. Кроме того, инсулин повышает проницаемость клеточных мембран для глюкозы. В связи с этим происходит усиленное проникновение глюкозы внутрь клеток, где осуществляется ее повышенное усвоение. Инсулин стимулирует синтез белка из аминокислот и активный транспорт их в клетки, задерживает распад белков и превращение их в глюкозу. Инсулин регулирует также жировой обмен — способствует образованию высших жирных кислот из продуктов углеводного обмена, а также тормозит мобилизацию жира из жировой ткани.
Рецепторы инсулина расположены на мембране клетки-мишени (рис. 10.5), поэтому первично гормон проявляет свое действие, не проникая в клетку. Связывание инсулина со специфическим рецептором клетки приводит к процессам, которые увеличивают скорость образования и накопления гликогена, белка и липидов. Активность инсулина выражается в лабораторных и клинических единицах. Лабораторная, или кроличья, единица — это то количество гормона, которое у здорового кролика массой в 2 кг уменьшает содержание глюкозы в крови до 2,22 ммоль/л (40 мг%). За одну единицу действия (ЕД) или интернациональную единицу (ИЕ) принимают активность 0,04082 мг кристаллического инсулина. Клиническая единица составляет 1/5 лабораторной.
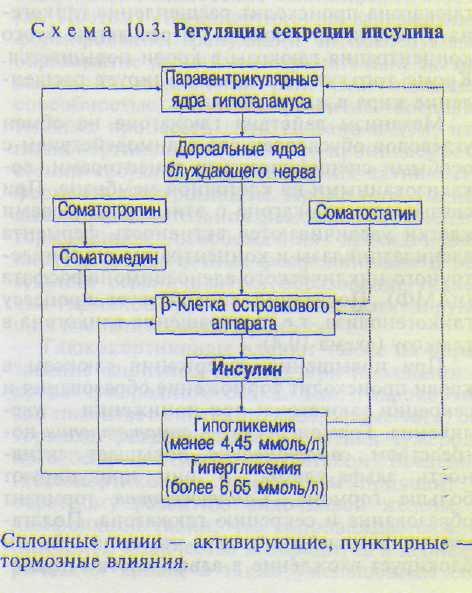
В основе регуляции образования и секреции инсулина лежит содержание глюкозы в крови. Гипергликемия приводит к увеличенному образованию и поступлению инсулина в кровь. Гипогликемия уменьшает образование и поступление гормона в сосудистое русло. Это осуществляется, во-первых, с помощью паравентрикулярных ядер гипоталами-ческой области. При увеличении концентрации глюкозы в крови происходит повышение активности нервных клеток паравентрику-лярного ядра. Возникшие в нейронах импульсы передаются к дорсальным ядрам блуждающего нерва, а по его волокнам — к бета-клеткам островков Лангерганса, и в них усиливаются образование и секреция инсу-
193

Рецептор/ \Мем6рзнные
\ / \эффекты
Инсулин у/ д ^^^
-Второй медиатор — Эффекты ^ ферментные
Инсулин+рецептор
. О Лизосома
-Клеточная мембрана
Рис. 10.5. Взаимодействие инсулина с рецептором (по Д.Бакстеру, Ф.Филингу, 1982).
лина (схема 10.3). Действие последнего снижает уровень глюкозы в крови. При снижении количества глюкозы в крови ниже нормы возникают противоположные реакции. Возбуждение симпатической нервной системы тормозит выделение инсулина.
Во-вторых, повышенный уровень глюкозы в крови возбуждает непосредственно рецеп-торный аппарат ткани поджелудочной железы, что также вызывает увеличение образования, секреции инсулина и снижение уровня глюкозы. При падении количества глюкозы в крови возникают противоположные реакции. Глюкоза стимулирует образование и секрецию инсулина также за счет непосредственного воздействия на бета-клетки островков Лангерганса. Полагают, что глюкоза взаимодействует с особым рецептором на мембране бета-клеток (глкжозорецептором), в резуль-
тате чего в них усиливаются синтез и освобождение инсулина в кровоток.
Секреция инсулина происходит и рефлек-торно при раздражении рецепторов ряда рефлексогенных зон. Так, при повышении уровня глюкозы в крови возбуждаются хеморе-цепторы каротидного синуса, в результате чего осуществляется рефлекторный выброс инсулина в кровоток и уровень глюкозы в крови восстанавливается. Стимулируют образование и секрецию инсулина также сома-тотропин аденогипофиза посредством сома-томединов, гормоны желудочно-кишечного тракта секретин и холецистокинин-панкреози-мин, а также простагландин Е за счет повышения аденилатциклазной активности мембран бета-клеток поджелудочной железы.
Соматостатин в противоположность со-матотропину тормозит образование и секрецию инсулина бета-клетками островков Лан-герганса. Он образуется в ядрах гипоталамуса и в дельта-клетках островковой части поджелудочной железы.
Количество инсулина в крови определяется также активностью фермента инсулиназы, который разрушает гормон. Наибольшее количество фермента содержится в печени и скелетных мышцах.
Глюкагон также принимает участие в регуляции углеводного обмена. По характеру своего действия на обмен углеводов он является антагонистом инсулина. Под влиянием глкжагона происходит расщепление гликогена в печени до глюкозы. В результате этого концентрация глюкозы в крови повышается. Кроме того, глюкагон стимулирует расщепление жира в жировой ткани.
Механизм действия глкжагона на обмен углеводов обусловлен его взаимодействием с особыми специфическими рецепторами, локализованными на клеточной мембране. При связывании глюкагона с этими рецепторами клетки увеличиваются активность фермента аде н ил ат цикл азы и концентрация внутриклеточного циклического аденозинмонофосфата (цАМФ). Последний способствует процессу гликогенолиза, т.е. превращения гликогена в глюкозу (схема 10.4).
При повышении содержания глюкозы в крови происходит торможение образования и секреции глюкагона, при понижении — увеличение. Гормон роста — соматотропин посредством соматомедина повышает активность альфа-клеток, и они продуцируют больше гормона. Соматостатин тормозит образование и секрецию глюкагона. Полагают, что это связано с тем, что соматостатин блокирует вхождение в альфа-клетки подже-
Схема 10.4. Регуляция секреции глюкагона
Соматостатин
Соматомедин
а-Клетки островкового аппарата