

9.4. Парасимпатическая нервная система
А. Иннервируемые органы и локализация пре-ганглионарных и ганглионарных нейронов.
Парасимпатические нервные волокна имеются в черепных нервах (III пара — мезэнце-фальный отдел, VII, IX и X пары — бульбар-ный отдел) и в тазовом нерве — сакральный отдел спинного мозга (S2—S4). Парасимпатические волокна Ш-й пары (глазодвигательный нерв) иннервируют глазные мышцы (т. sphincter pupillae и т. ciliaris), регулируя диаметр зрачка и степень аккомодации. Парасимпатические веточки VII пары (лицевой нерв): п. petrosus major — секреторный нерв, иннервирует слизистую оболочку носа, неба, слезную железу; п. chorda tympani — смешанный нерв, содержит чувствительные и секреторные волокна подчелюстной и подъязычной слюнных желез. Парасимпатические секреторные волокна IX пары (языкоглоточный нерв) подходят к околоушной железе в составе п. auriculotemporalis — от третьей ветви тройничного нерва. X пара (блуждающий нерв) своими ветвями снабжает дыхательные органы, большую часть пищеварительного тракта (до нисходящей ободочной кишки), сердце, печень, поджелудочную железу, почки. Парасимпатические нервы сакрального отдела спинного мозга (S2—-S4) иннервиру-
ют нисходящую часть ободочной кишки и тазовые органы (прямую кишку, мочевой пузырь, половые органы). Парасимпатической иннервации не имеют скелетные мышцы, матка, мозг, подавляющее большинство кровеносных сосудов (кожи, органов брюшной полости, мышц), органы чувств и мозговое вещество надпочечников.
Б. Парасимпатические ганглии и отдельные нейроны расположены внутри органов, а в тазовой области и в области головы — в непосредственной близости от органов. От нервных клеток парасимпатических ганглиев идут короткие постганглионарные парасимпатические волокна, иннервирующие все перечисленные органы; преганглионарные волокна обычно длинные (у симпатической нервной системы, наоборот, преганглионарные — короткие, постганглионарные — длинные).
В. Медиаторы и рецепторы.
-
Передача возбуждения с преганглиопар- кого парасимпатического волокна на эффек- торный нейрон осуществляется, как и у сим патического отдела ВНС, с помощью ацетил- холина. Медиатор действует на Н-холиноре- цепторы постсинаптической мембраны ган- глионарного нейрона.
-
Постганглионарное волокно свое влия ние на эффекторную клетку передает также с помощью ацетилхолина, который в парасим патических терминалях находится в трех фондах (пулах): 1) стабильном, прочно свя занном с белком, не готовом к освобожде нию; 2) мобилизационном, менее прочно связанном и пригодном к освобождению; 3) готовом к освобождению ацетилхолина (активный медиатор), который освобождает ся квантами спонтанно и при поступлении к нервному окончанию потенциалов действия. Активный медиатор находится в прилежащих к мембране пузырьках. Пузырьки с медиато ром находятся в движении, и по мере расхо дования активного медиатора к пресинапти- ческой мембране поступают новые пузырьки с активным ацетилхолином. Освобождению квантов медиатора способствуют ионы Са3+. Ацетилхолин синтезируется в цитоплазме окончаний холинергических нейронов, депо нируется в везикулах по нескольку тысяч мо лекул в каждой из них.
-
Инактивирование медиатора. Выделив шийся в синаптическую щель ацетилхолин, как и в любом другом синапсе, не весь ис пользуется для передачи сигнала. В отличие от симпатической нервной системы основная часть ацетилхолина разрушается ферментом ацетилхолинэстеразой с образованием холи-
164
на и уксусной кислоты, которые захватываются пресинаптической мембраной и вновь используются для синтеза ацетилхолина. Значительно меньшая часть медиатора диффундирует в интерстиций и кровь. Обратного захвата нерасщепленного ацетилхолина нервными окончаниями не происходит. По поводу локализации ацетилхолинэстеразы существует две точки зрения: согласно одной из них, фермент фиксирован на постсинапти-ческой мембране вблизи холинорецепторов, согласно другой, — на базальной мембране. Последняя представляет собой тонкую сеть коллагена и гликозаминогликанов между пре- и постсинаптическими мембранами.
4. Эффекшорные рецепторы. На клетки-эффекторы ацетилхолин действует с помощью М-холинорецепторов (см. рис. 9.2), которые свое название получили от мускари-на — токсина мухомора, активирующего эти рецепторы и вызывающего такой же эффект, как и ацетилхолин. Мускариноподобный эффект ацетилхолина был открыт позже. М-хо-линорецепторы имеются также на симпатических и парасимпатических окончаниях, в коре головного мозга, ретикулярной формации. По чувствительности к различным фармакологическим препаратам выделяют М,— М4-холинорецепторы. М,-холинорецепторы локализуются в вегетативных ганглиях и ЦНС, Мгхолинорецепторы — в сердце, на пресинаптических окончаниях, в гладких мышцах желудочно-кишечного тракта. В гладких мышцах расположены также М3-хо-линорецепторы, имеются они и в большинстве экзокринных желез. Мгхолинорецепторы изучены мало.
Некоторые симпатические нервные волокна (иннервирующие потовые железы и вызывающие расширение сосудов скелетных мышц) также являются холинергическими.
Убедительного объяснения этому факту до настоящего времени не имеется. Сам по себе факт передачи сигнала с постганглионарных симпатических волокон посредством ацетилхолина считается установленным. Однако постганглионарное симпатическое волокно является аксоном адренергического нейрона, в его окончаниях синтезируется норадрена-лин, что противоречит сложившемуся представлению о «холинергических» симпатических нервных волокнах. Согласно гипотезе Я.А. Росина, потовые железы иннервируются преганглионарными симпатическими волокнами, которые, как известно, выделяют ацетилхолин. Возможно также, что потовые железы иннервируются симпатическими по-стганглионарными волокнами, реализующими свое стимулирующее на потоотделение влияние с помощью норадреналина и активации р-адренорецепторов.
Г. Связь постганглионарных парасимпатических окончаний с холинорецепторами более тесная, нежели у симпатических окончаний. Короткие постганглионарные волокна холинергических нейронов мало ветвятся и образуют типичные синапсы с клетками-эффекторами. Однако имеются и неиннервирован-ные постсинаптические М-холинорецепто-ры, например, в кровеносных сосудах.
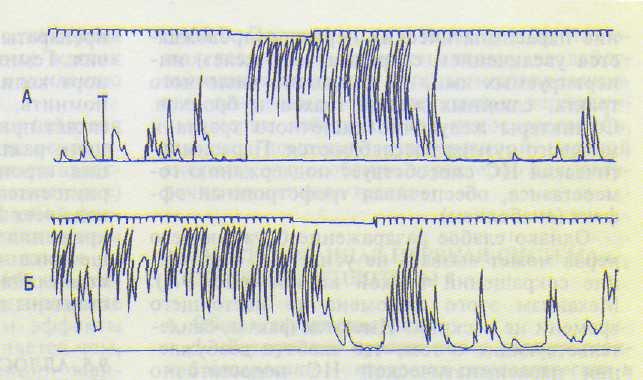
Д. Эффекты активации М-холинорецепторов приводят к сокращению гладкой мускулатуры желудочно-кишечного тракта — перистальтика усиливается (рис. 9.6, А), к сокращению мускулатуры бронхов — их просвет сужается, к сокращению мышц мочевого пузыря, сокращению сфинктера зрачка — зрачок сужается, сокращению ресничной мышцы глаза — хрусталик становится более выпуклым. Одновременно тормозится деятельность сердца, расширяются сосуды половых и некоторых других органов. Возбужде-
165
ние парасимпатических нервов сопровождается увеличением секреции всех желез, ин-нервируемых ими, — желудочно-кишечного тракта, слюнных желез, трахеи и бронхов. Сфинктеры желудочно-кишечного тракта и мочевого пузыря расслабляются. Парасимпатическая НС способствует поддержанию го-меостазиса, обеспечивая трофотропный эффект (анаболизм).
Однако слабое раздражение блуждающего нерва может вызвать не усиление, а угнетение сокращений тонкой кишки (рис. 9.6). Механизм этого феномена до настоящего времени не раскрыт. Имеются факты, свидетельствующие о том, что слабого возбуждения парасимпатической НС недостаточно для возбуждения гладких мышц, но достаточно для активации адренергических нервных окончаний, возбуждение которых и ведет к угнетению сокращений кишки.
Эффекты экзогенного ацетилхолина те же, что и парасимпатической нервной системы. Однако в клинической практике ацетилхолин не применяется, так как очень быстро разрушается. С лечебной целью применяется более стойкий препарат — карбохолин, он не разрушается холинэстеразой.
Е- Механизм действия ацетилхолина и чувствительность эффекторных клеток к медиатору. Стимулирующее влияние ацетилхолина на орган осуществляется, во-первых, посредством изменения характера электрофизиологических процессов — возбуждения посредством активации ионотропных рецепторов Ыа+-каналов; во-вторых — посредством активации метаботропных рецепторов и соответствующих биохимических реакций с помощью вторых посредников: инозитолтрифос-фата, Са . Тормозный эффект ацетилхолина возникает также в результате изменения характера электрофизиологических процессов (активации ионотропных рецепторов К+-ка-налов и гиперполяризации клеток эффектора). При этом с помощью метаботропных рецепторов активируется система вторых посредников: гуанилатциклаза-циклический гу-анозинмонофосфат (ГЦ-цГМФ), обладающая анаболическим эффектом. Подобное действие выражено также у брадикинина, гистамина, инсулина. Однако гистамин посредством Н2-рецепторов может активировать и аденилатциклазную систему. Атропин и скополамин блокируют и возбуждающие, и тормозящие холинорецепторы, а значит, и эффекты ацетилхолина, что широко применяется в клинической практике с лечебной и диагностической целью. С целью блокады парасимпатических эффектов применяются
препараты и с другими механизмами действия. Гемихолин, например, нарушает транспорт холина, тормозит его синтез. Следует помнить, что блокада любых рецепторов может привести к разнонаправленным реакциям различных органов. Например, инъекция атропина, который, попадая в кровь, разносится по всему организму, ведет к блокаде всех М-холинорецепторов, при этом сокращения сердца учащаются, а желудка и кишечника — тормозятся. Основные эффекты стимуляции парасимпатических нервов представлены в табл. 9.1.
9.5. АЛЛОСТЕРИЧЕСКОЕ
(ЭЛЕКТРОФИЗИОЛОГИЧЕСКОЕ)
И АЛЛОХРОННОЕ (БИОХИМИЧЕСКОЕ)
ДЕЙСТВИЕ МЕДИАТОРА
Терминология. Электрофизиологическое действие медиатора предлагают называть «аллостеричес-ким», а биохимическое — «аллохронным». При этом под аллостерическим действием медиатора понимают совокупность процессов взаимодействия медиатора с эффекторными рецепторами на поверхности клеточной мембраны, в результате чего возникают электрические ответы вследствие активации ионных каналов и движения различных ионов в клетку и из клетки. Действие медиатора на биохимические реакции, в том числе на процессы клеточного синтеза и считывания информации с молекул ДНК, называют аллохронным. Аллохронное (греч. allos — другой + chroni-kos — относящийся ко времени) и аллостеричес-кое (stereos — пространственный) действие медиатора — термины, отражающие эффект действия медиатора, которые следует уточнить. Во-первых, оба процесса протекают во времени, во-вторых, в обоих случаях протекают те или иные динамические процессы: изменяется проницаемость клеточной мембраны, взаимодействуют молекулы различных веществ. Поэтому термин «электрофизиологическое» действие медиатора нецелесообразно заменять на «аллостерическое», а «биохимическое» — на «аллохронное».
Поскольку электрофизиологическое действие медиаторов (оно реализуется с помощью ионотропных рецепторов) выражается в одних случаях в гиперполяризации клеточных мембран, в других — в деполяризации, один и тот же нерв с помощью одного и того же медиатора в разных клетках может вызывать различные эффекты. Так, возбуждение симпатического нерва вызывает возбуждение кард и ом и о пито в и усиление деятельности сердца, а деятельность глад ком ышечных клеток обычно угнетается вследствие их гиперполяризации. Медиатор блуждающего нерва, наоборот, вызывает гиперполяризацию кар-
166
диомиоцитов, угнетение деятельности сердца и деполяризацию гладкомышечных клеток желудка и кишечника, что ведет к усилению их деятельности.
Основные схемы биохимического действия медиатора. Одним из механизмов реализации химического эффекта (он осуществляется с помощью метаботропных рецепторов) является активация аденилатциклазной системы: связывание медиатора рецепторным белком — активация аденилатциклазы — продукция молекул цАМФ из АМФ — активация протеинкиназ — фосфорилирование фермента. Вызываемые при этом эффекты весьма разнообразны. Это объясняется тем, что характер биохимических реакций зависит не от особенностей строения цАМФ, а от специфичности реагирующих клеток. При этом имеет место каскад усилений эффекта медиатора или гормона: молекула медиатора — рецепторный белок — несколько аденилатциклазных комплексов, каждый из которых обеспечивает продукцию нескольких тысяч молекул цАМФ и активацию еще большего числа молекул протеинкиназ. цАМФ как второй посредник изучен лучше цГМФ, кальция, производных ф°сФатиДи-иинозитола.
цГМФ (циклический гуанозинмонофос-фат) локализуется, как и цАМФ, на клеточной мембране. Схема его действия та же, что и у цАМФ: медиатор — рецептор клеточной мембраны — активация гуанилатциклазы — продукция цГМФ из ГТФ — действие цГМФ — активация протеинкиназ — фосфорилирование фермента — субстрат — эффект. Концентрация цГМФ в тканях в 10—15 раз меньше, нежели цАМФ, уже и набор протеинкиназ, нередко эффекты цГМФ и цАМФ противоположны. Уровень цГМФ в тканях повышается под влиянием ряда гормонов и медиаторов: секретина, инсулина, гистамина, окси-тошша, серотонина, брадикинина, ацетилхо-лина. При этом активируются анаболические процессы, подобно влияниям парасимпатической нервной системы. В свою очередь эр-готропное действие (катаболический эффект) оказывают не только норадреналин и адреналин, но и другие вещества, связанные с активностью симпатической нервной системы, в частности дофамин, октопамин, серотонин, АКТГ, ТТГ, вазопрессин. Однако у них могут быть и другие эффекты. Сам адреналин может вызвать двоякий метаболический эффект — посредством а2-адренорецепторов он способен ингибировать аденилатциклазу, а посредством р-адренорецепторов активирует аденилатциклазу, что ведет к накоплению
цАМФ. Поэтому одна и та же клетка, имеющая различные адренорецепторы, на действие одного и того же лиганда может реагировать по-разному. Экстраорганные вегетативные нервы действуют как непосредственно, так и с помощью интраорганной нервной системы, что особенно характерно для желудочно-кишечного тракта.