

9.2. Симпатическая нервная система
Симпатическая нервная система характеризуется рядом особенностей.
А. Локализация преганглионарных и ган-глиоыарных нейронов и иннервируемые ими органы. В отличие от парасимпатических нервов, которые берут начало из разных отделов ЦНС, все симпатические нервы выходят из спинного мозга (нейроны расположены в боковых рогах, сегменты СУщ—L,, — центр Якобсона) и иннервируют все органы и ткани (рис. 9.1). В последнем шейном и двух первых грудных сегментах спинного мозга расположен спиноцилиарный центр, иннервирующий гладкие мышцы глаза, в том числе мышцу, расширяющую зрачок. Эффек-торные нейроны этого пути лежат в краниальном шейном симпатическом ганглии. От 2—4 грудных сегментов спинного мозга отходят симпатические волокна, иннервирующие слюнные железы. В первых пяти грудных сегментах спинного мозга расположены эффек-торные симпатические нейроны, иннервирующие сердце. Ганглионарные клетки этого пути лежат в основном в звездчатом ганглии или, реже, — в узлах пограничного симпатического ствола. Сосуды и потовые железы иннервируются от всех симпатических преганглионарных нейронов спинного мозга.
Из спинного мозга преганглионарные симпатические волокна выходят в составе передних корешков и, отделившись от них, идут в виде белых соединительных ветвей к симпатическому стволу. Это преганглионарные, мя-котные (миелиновые) волокна, часть из которых прерывается в узлах симпатического ствола, другая часть проходит не прерываясь.
Выходят из симпатического ствола два нервных пути. Первый из них — серые соединительные ветви, представляющие собой постганглионарные симпатические волокна и вступающие в спинномозговые (соматические) нервы. Они иннервируют лимфатические и кровеносные сосуды, железы, мышцы, поднимающие волосы туловища и конечностей, скелетную мускулатуру, все органы и ткани. Второй путь — нервы, идущие непосредственно к внутренним органам и сосудам. Небольшая часть волокон этих нервов является постганглионарными, большая — преганглионарными. Последние образуют синапсы на клетках превертебральных ганглиев, а также шейных, нижних поясничных, крестцовых и копчиковых. От нейронов этих ганглиев идут постганглионарные волокна, иннервирующие внутренние органы и ткани организма. Большинство симпатических ган-
глиев удалено от иннервируемых ими органов, поэтому от этих ганглиев идут довольно длинные постганглионарные аксоны. Исключение составляют лишь некоторые относительно небольшие симпатические ганглии, расположенные рядом с половыми органами и посылающие к ним короткие постганглионарные волокна.
Таким образом, симпатические нервы регулируют функции всех органов и тканей организма, включая ЦНС и сенсорные рецепторы. Об этом же свидетельствуют и следующие данные.
Норадренергические нейроны голубого пятна по своим морфологическим, биохимическим и электрофизиологическим свойствам весьма сходны с периферическими нор-адренергическими нервными клетками. Есть данные о том, что многие норадренергические волокна, исходящие от нижних частей ствола мозга, иннервируют артериолы и капилляры коры большого мозга. Подобные нейроны могут участвовать в регуляции кровотока через кору, и их можно рассматривать как центральный отдел симпатической нервной системы.
Б. Медиаторы и рецепторы. 1. Преганглионарные нейроны и рецепторы ганглионарных нейронов. Эфферентный вход в вегетативном ганглии (экстра- и интраорганный) представлен возбуждающим холинергическим преган-глионарным волокном, образующим синапс с ганглионарным нейроном с помощью Н-холинорецептора (медиатор — ацетилхо-лин). Рецептор получил свое название (Д. Ленгли) из-за чувствительности к никотину: малые его дозы возбуждают нейроны ганглия, большие блокируют. В вегетативных ганглиях, кроме ацетилхолина, имеются ней-ро пептиды: метэнкефалин, нейротензин, ВИП, холецистокинин, вещество П, но их роль как медиаторов не доказана. Н-холино-рецепторы локализованы также на клетках скелетных мышц, каротидных клубочков и мозгового слоя надпочечников. Н-холиноре-цепторы нервно-мышечных соединений и вегетативных ганглиев блокируются различными фармакологическими препаратами. Например, бензогексоний блокирует Н-хо-линорецепторы вегетативных ганглиев, но не блокирует Н-холинорецепторы нервно-мышечного соединения. Последние блокируются ядом кураре и другими препаратами, которые не блокируют Н-холинорецепторы вегетативных ганглиев. В ганглиях обнаружены также М-холинорецепторы (возбуждаются мускарином) и опиатные рецепторы. Они регулируют выделение медиатора — ацетилхо-
158
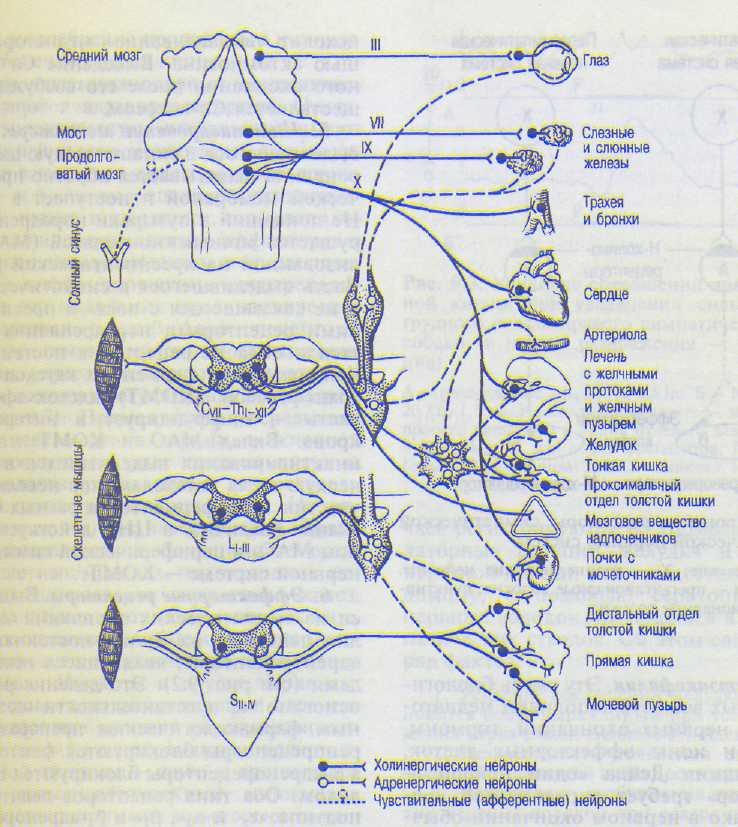
Рис. 9.1. Холинергическая и адренергическая иннервация. С, Th, L, S, — сегменты спинного мозга.
лина в синапсах ганглия и чувствительность Н-холинорецепторов. В нейронах ганглиев можно зарегистрировать тормозные — гиперполяризационные потенциалы. Вызываются они вставочными адренергическими клетками, регулирующими возбудимость ганглио-нарных клеток.
2. От ганглионарных нейронов отходят по-апганглионарные симпатические волокна, в окончаниях которых главным медиатором является норадреналин — около 90 % (адреналин — около 7 %, дофамин — около 3 %). Синтезируется норадреналин из аминокислоты тирозина в аксоне ганглионарного нейрона, в основном в терминальных симпатических окончаниях, около 1 % — в теле нейрона. Поскольку в окончаниях симпатических ганглионарных нейронов выделяется норадрена-
лин, эти нейроны называют адренергическими (рис. 9.2). Из симпатических нервных окончаний выделяется не только медиатор норадреналин, но и комедиатор нейропептид Y, тогда как из парасимпатических окончаний наряду с ацетилхолином выделяется ва-зоактивный интестинальный пептид. Необходимо отметить, что все нервные окончания имеют два основных типа секреторных везикул. Мелкие синаптические везикулы имеют диаметр около 50 нм, однородны по размерам и содержат классические медиаторы. Крупные, электронно-плотные везикулы имеют диаметр около 100 нм, неоднородны по размерам и содержат пептиды.
3. В ответ на раздражение симпатического нерва вместе с норадреналином выделяются также белок хромагринин, дофамин-$-гид-
159

а- и р- адренорецепторы М-холинорецептор
Рис. 9.2. Нейроны и рецепторы симпатической и парасимпатической нервных систем.
А — адренергические, X — холинергические нейроны. Сплошная линия — прегангл ион арные волокна; пунктирная — постганглионарные волокна.
роксилаза, метэнкефалин. Эту смесь биологически активных веществ дополняют медиаторы соседних нервных окончаний, гормоны, метаболиты и ионы эффекторных клеток, поэтому принцип Дейла «один нейрон — один медиатор» требует определенной коррекции. Однако в нервном окончании обычно имеется основной медиатор. В симпатических нервных окончаниях основным медиатором является норадреналин. Он находится в гранулах в двух видах: резервном (стабильном) — 85—90 % и мобильном, который мобилизуется при поступлении нервных импульсов и выделяется из симпатических тер-миналей. Мобильный норадреналин пополняется из стабильного фонда.
4. Наиболее значим в процессах выделения медиатора при возбуждении нервного окончания везикулярный экзоцитоз, состоящий из нескольких этапов. Первый этап (сопряжение деполяризации и секреции) реализуется с помощью Са2+, который проникает в пре-синаптические нервные окончания через медленные и быстрые Са-каналы. На место выделившихся везикул медиатора транспортируются по филаментам с затратой энергии новые гранулы медиатора. Источник энергии — АТФ. Когда Са2+обеспечивает попада-
ние мелмяторя и
160
исходит «выдавливание» медиатора с помощью актомиозина. Выведение Са2+ из нервного окончания после его возбуждения осуществляется Са-насосом.
-
Инактивирование медиатора. 75—80 % выделившегося в синаптическую щель норад- реналина захватывается обратно пресинапти- ческой мембраной и поступает в пузырьки. Не попавший в пузырьки норадреналин раз рушается моноаминоксидазой (МАО), лока лизованной на пресинаптической мембране. Часть выделившегося в синаптическую щель и не связавшегося с пост- и пресинаптичес- кими рецепторами норадреналина разруша ется в области рецепторов постсинаптичес- кой мембраны ферментом катехол-О-метил- трансферазой (КОМТ) клеток-эффекторов, часть — диффундирует в интерстиций и кровь. Вклад МАО и КОМТ в процессы инактивирования выделившихся в синапти ческую щель катехоламинов невелик, а доля участия этих ферментов в разных органах и тканях различна: в ЦНС действует в основ ном МАО, в периферической симпатической нервной системе — КОМТ.
-
Эффекторные рецепторы. Выделивший ся из симпатических окончаний норадрена лин действует на а- и р-постсинаптические адренорецепторы, являющиеся гликопротеи- дами (см. рис. 9.2). Это деление рецепторов основано на чувствительности их к различ ным фармакологическим препаратам: а-ад- ренорецепторы блокируются фентоламином, а р-адренорецепторы блокируются пропрано- лолом. Оба типа рецепторов делятся на два подтипа: а,- и а2-, рг и р2-адренорецепторы. Антагонистами а,-адренорецепторов являют ся празозин, дроперидол, антагонисты сс2-ад- ренорецепторов — раувольсин, йохимбин. Антагонистами р,-адренорецепторов явля ются практолол, атенолол, р2-адренорецепто- ров — бутоксамин.
-
Распределение а~ и $-адренорецепторов в различных органах. В большинстве органов, реагирующих на катехоламины, содержатся а- и р-адренорецепторы, причем одна гладкомы- шечная клетка может иметь оба рецептора (см. табл. 9.1). Распределение а- и р-адрено- рецепторов в сосудах организма подробнее см. раздел 13.9.1. На тромбоцитах имеются аг адренорецепторы, стимуляция которых повы шает их агрегацию, и р2-адренорецепторы, выполняющие противоположную функцию. В чистом виде выделены из мембраны агад- ренорецепторы: они представляют собой бе лок с молекулярной массой 9600 и обусловли вают сокращение сосудов, матки, СШЯ$Ш9~
протоков, семенных пузырьков, пило-
моторов и расслабление продольного и циркулярного мышечных слоев кишечника.
В. Связь постганглионарных симпатических окончаний с адренорецепторами. Считают, что симпатическую иннервацию имеют только а,- и р|-адренорецепторы. Активируются они в основном медиатором норадрена-лином. а2- и Р2-Адренорецепторы преимущественно не имеют иннервации, они расположены вне синапсов на клетках-эффекторах и активируются циркулирующим в крови адреналином и диффундирующим норадренали-ном после его выделения пресинаптически-ми окончаниями. а2- и р2-АдРеноРеиепторы обнаружены также на пресинаптических окончаниях (пресинаптические рецепторы), где они выполняют регулирующую функцию (см. раздел 9.7). Прямая связь посредством синапса имеет место в сердце, в жировой ткани, в сосудах и в ряде гладкомышечных органов (мышцы зрачка, матки, пиломоторы, семявыносящий проток, кишечник). Непрямой тип регуляции, как известно (В.А. Говы-рин), осуществляется в большинстве органов — скелетных мышцах, эндокринных железах, большинстве экзокринных желез. В этих органах медиатор из сосудистых симпатических сплетений диффундирует к местным окружающим сосуд клеткам либо разносится кровью. Имеются и другие варианты: симпатические (постганглионарные адренер-гические) нервные волокна образуют синап-тические контакты с нейронами интраорган-ной нервной системы — либо с преганглио-нарными парасимпатическими волокнами, либо с ганглионарными холинергическими нейронами посредством а-рецепторов. Подобные взаимодействия симпатической и парасимпатической нервных систем установлены в органах желудочно-кишечного тракта, предполагаются в других органах.
Г. Эффекты активации а- и р-адренорецеп-торов. В физиологических условиях реакция какого-либо органа на адреналин и норадре-налин, поступающие с кровью либо выделяющиеся при возбуждении симпатических нервов, зависит от преобладания а- или р-ад-ренорецепторов в данном органе и различной их чувствительности.
Активация си- и а2-адренорецепторов мембран клеток-эффекторов ведет к деполяризации их и к повышению активности органа, в том числе сфинктеров желудочно-кишечного тракта. Однако у миоцитов желудка и кишечника развивается гиперполяризация, при этом мышечная стенка органов расслабляется.
Если же симпатический ствол раздражается в грудной полости, то в большинстве слу-

Рнс. 9.3. Усиление сокращений двенадцатиперстной кишки при увеличении силы раздражения грудного отдела правого симпатического ствола у собаки (в начале раздражения — слабое угнетение).
А — раздражение 5 В, 20 Гц, 0,5 мс. Б — раздражение 10 В, 20 Гц, 1,5 мс. На каждом фрагменте — запись давления в полости кишки (I) и линия нулевого давления (2); шкала 0—20 мм рт.ст. для обоих фрагментов; Р — раздражение (опыт В.М-Смирнова, Д.С.Свешникова, И.Л. Мясникова).
чаев регистрируются не тормозные, а стиму-ляторные реакции желудка и двенадцатиперстной кишки. Это осуществляется, по-видимому, с помощью серотонинергических нервных волокон, имеющихся в составе симпатических стволов. Об этом свидетельствует ряд фактов.
-
Усиление сокращений желудка и кишки ста новится более выраженным при усилении раздра жения нерва (закон силовых отношений) (рис. 9.3).
-
Независимость появления стимуляторного эффекта от фоновой активности органа.
-
Исключение возбуждения парасимпатичес ких нервных волокон при раздражении симпати ческого нерва не влияет на степень выраженности стимуляторного эффекта.
-
Блокада различными фармакологическими препаратами адренергических окончаний, а- и Р- адренорецепторов или Н-холинорецепторов веге тативных ганглиев не только не устраняет, а, на против, усиливает стимуляторный эффект раздра жения симпатического нерва.
-
Блокада S3 (5-НТз)-серотонинорецепторов вегетативных ганглиев и Si)2 (5-НТи)-серотони- норецепторов гладкой мышцы желудка и кишки устраняет стимуляторный эффект раздражения симпатического нерва.
-
Блокада серотонинорецепторов вегетатив ных ганглиев и гладкой мускулатуры не влияет на функции симпатического и парасимпатического нервов (точнее, адренергического и холинерги- ческого отделов ВНС).
-
В составе грудного отдела симпатического ствола обнаружен серотонин с помощью гистохи мической и биохимической методик. Стимулятор- ные эффекты желудка и тонкой кишки впервые получили с помощью раздражения симпатическо го ствола в грудной полости у собак, где он не со держит парасимпатических волокон.
161

Рис. 9.4. Функциональная организация эфферентного отдела вегетативной нервной системы (на примере желудка и тонкой кишки.
Нейроны: А — адреН-, С — серотонин-, X — ХОЛИИ-, П — пуринергические и соответствующие им а-, р-, S-, Н-, М-и Р-рецепторы. Сплошная линия — преганглионарные, пунктир — постганглионарные волокна.
Общая схема эфферентной иннервации желудка и тонкой кишки, ганглионарные и эффекторньге рецепторы представлены на рис. 9.4.
Активация $-адренорецепторов вызывает разнонаправленные эффекты в различных органах. Активация (3-адренорецепторов в
сердце (ргрецепторы хорошо иннервирова-ны) ведет к деполяризации и возбуждению кардиомиоцитов, к увеличению частоты и силы сердечных сокращений, к повышению автоматизма, облегчению атриовентрикуляр-ной проводимости. При активации руадре-норецепторов эффекторных клеток гладко-мышечных органов (р2-адренорецепторы ин-нервированы меньше) возникает гиперполяризация, наблюдается угнетение функции органа, в частности расширение сосудов. При одновременной активации а- и $-адренорецеп-торов сосуды суживаются вследствие преобладания а-адренорецепторов (рис. 9.5).
Д. Механизм действия катехоламинов и чувствительность эффекторных клеток к медиатору.
1. Во всех видах адренорецепторов кате-холамины взаимодействуют посредством G-белка. При этом возникают электрофизиологические процессы (деполяризация или гиперполяризация в результате активации ио-нотропных рецепторов). При деполяризации наблюдается усиление функции органа (например, усиление сокращений сердца), при гиперполяризации — угнетение (например, уменьшение тонуса кишки). При активации метаботропных рецепторов возникают метаболические сдвиги (биохимические процессы) с помощью вторых посредников: при активации а-адренорецепторов — инозитол-3-фосфата (ИФ3) и Са2+; при активации р-адре-норецепторов — аденилатциклазы-цАМФ. Мессенджерами для катехоламинов являются также цГМФ, диацилглицерин (ДАГ).

А — интактные а- и В-адренорецепторы; Б — блокада а- и В-адренорецепторов; В -блокада а-адренорецепторов.
G-белок важен для функционирования и других рецепторов. Более 80 типов рецепторов связано с ионными каналами или ферментами посредством G-белка.
2. Чувствительность эффекторных клеток к медиатору может понижаться (десен-ситизация) или повышаться (сенситизация). Эти явления достаточно хорошо изучены на примере регуляции деятельности сердца симпатической нервной системой. В основе де-сенситизации при избытке медиатора лежит снижение общего числа рецепторов в результате их разобщения с каталитической субъединицей и последующего эндоцитоза рецепторов. Напротив, сенситизация является следствием увеличения числа рецепторов на эффекторных клетках и уменьшения количества ферментов, разрушающих тот или иной медиатор, выделяющийся из пресинаптичес-кого окончания. Это наблюдается, например, в эксперименте при перерезке нерва или блокаде выделения медиатора из нервного окончания фармакологическим препаратом. В клинической практике с лечебной целью нередко блокируют или стимулируют постси-наптические рецепторы, процессы синтеза, высвобождения и разрушения медиаторов, что сопровождается изменением чувствительности эффекторных клеток к гуморальным и нервным влияниям. В частности, в условиях блокады выделения медиатора нервными окончаниями или истощения его запасов в нервных элементах, например, с целью снижения артериального давления развивается сенситизация за счет увеличения числа рецепторов на эффекторных клетках. Поэтому после отмены подобного препарата артериальное давление снова повышается, причем в большей степени, что необходимо учитывать в клинической практике. Число адреноре-цепторов на эффекторной кле-гке увеличивается под влиянием гормонов щитовидной железы.
9.3. МОЗГОВОЙ СЛОЙ НАДПОЧЕЧНИКОВ
Мозговой слой надпочечников представляет собой видоизмененный симпатический ганглий — его клетки, с онтогенетической точки зрения гомологичны ганглионарным адре-нергическим нейронам. Они содержат включения, окрашивающиеся в желто-коричневый цвет двухромовокислым калием, что и послужило поводом назвать их хромаффин-ными клетками. В виде скоплений хромаф-финные клетки встречаются также на поверхности аорты, в области каротидного си-
нуса, среди клеток симпатических ганглиев. Преганглионарные волокна образуют на этих клетках, как и на хромаффинных клетках надпочечников, возбуждающие холинерги-ческие синапсы. Выделение катехоламинов из мозгового вещества надпочечников регулируется исключительно нервными влияниями (перерезка преганглионарных симпатических волокон прекращает секрецию катехоламинов). При возбуждении преганглионарных симпатических волокон у человека из надпочечников в кровоток обычно выбрасывается смесь катехоламинов, состоящая из адреналина (80—90 %) и норадреналина (10— 20 %). Точки приложения для продуцируемых надпочечниками катехоламинов те же, что и у симпатической нервной системы, однако их действие более выражено, нежели симпатических нервов в областях со слабой адренергической иннервацией (в круговых и продольных мышцах кишечника, крупных артериях, матке). Взаимодействие катехоламинов с адренорецепторами вызывает различные эффекты в разных органах, в частности торможение деятельности желудочно-кишечного тракта, улучшение процесса передачи в нервно-мышечных синапсах, увеличение силы сокращений скелетных мышц, частоты и силы сокращений сердца, расширение бронхов. Все это имеет важное приспособительное значение, обеспечивая мобилизацию систем организма при физическом и эмоциональном напряжениях.
Катехоламины усиливают высвобождение свободных жирных кислот из подкожной жировой ткани и образование из гликогена глюкозы, необходимой клеткам организма при состояниях напряжении. Ускорение расщепления углеводов осуществляется с помощью активации аденилатциклазы, стимулирующей образование цАМФ, который в свою очередь активирует фосфорилазу, расщепляющую гликоген — источник энергии. Таким образом, катехоламины мозгового слоя надпочечников можно рассматривать как метаболические гормоны. Не случайно симпатэктомированные животные не в состоянии осуществлять физические усилия, плохо переносят охлаждение и перегревание, с большим трудом справляются с кровотечением. У симпатэктомированных животных не бывает проявления характерных защитных реакций и показателей агрессивности: тахикардии, повышения артериального давления, расширения зрачков. Введение этим животным ганглиозидов ускоряет рост новых синапсов, ускоряет реиннервацию после повреждения нервных волокон. Клетки, подоб-
163
ные хромаффинным мозгового слоя надпочечников, называют трансдукторами, их короткие аксоны не имеют синаптических контактов с другими клетками, они выделяют свои биологически активные вещества, называемые также гормонами, в кровь. К клет-кам-трансдукторам относят, кроме хромаф-финных клеток, нейроны супраоптического и паравентрикулярного ядер гипоталамуса, выделяющие вазопрессин и окситоцин; нейроны гипоталамуса, выделяющие в сосудистую систему нейрогормоны, регулирующие функцию гипофиза (см. раздел 3.2), клетки юкстагломерулярного аппарата почки, которые под влиянием постганглионарных симпатических волокон выделяют в кровь ренин (см. разделы 13.9.2; 17.5.2).
Таким образом, симпатико-адреналовая система активирует деятельность организма, мобилизует его защитные силы, обеспечивает выход крови из кровяных депо, поступление в кровь глюкозы, ферментов, усиливает метаболизм тканей, увеличивает расход энергии, ее возбуждение является пусковым звеном стрес-сорных эмоционально-окрашенных реакций.