

Эпохи великих вымираний
В главе 2 уже говорилось о галактических циклах разной продолжительности, которым подчинены различные события земной истории, в том числе вымирание и возникновение организмов. Эта точка зрения развивается в последние годы академиком Е.Е.Милановским. По его мнению, периоды вымирания организмов являются следствием глобальных этапов тектонической активности земной коры.
Е.Е.Милановский (1999) предполагает, что взаимосвязанные, циклически проявлявшиеся изменения в развитии тектонических движений, магматизма, условий седиментации, морфогенеза на поверхности суши и ложе морей и океанов, в составе атмосферы и гидросферы, положении уровня Мирового океана могли оказывать значительное влияние на эволюцию органического мира Земли, на массу живого вещества биосферы, на таксономическое разнообразие морской подводной биоты. Все эти циклические изменения условий существования жизни на Земле, вероятно, в той или иной мере влияли на неоднократно возникавшие биотические кризисы и вызывли массовые, в том числе наиболее катастрофические среди них - так называемые великие вымирания значительной части обитателей нашей планеты.
В последнее время были достигнуты большие успехи в изучении изменений таксономического разнообразия биоты в фанерозое и феномена ее вымираний. Установлено, что на рубеже венда и кембрия произошло практически полное вымирание "эдиакарской" фауны многоклеточных мягкотелых бесскелетных животных. Затем в раннем кембрии появились и вскоре получили весьма широкое развитие многие группы морских организмов со скелетными элементами, но значительная их часть (более 40% родов и 25% семейств) подверглась быстрому вымиранию в середине и конце кембрия. В течение ордовика таксономическое разнообразие морской биоты вновь быстро возрастало, превысив к концу этого периода соответствующие показатели для конца кембрия в 3-4 раза. На рубеже ордовика и силура произошло новое, очень быстрое и значительное вымирание,
когда исчезло около 35% всех семейств, более 50% родов и около 65% видов всех ордовикских морских организмов.
Менее значительное, но все же существенное вымирание части морской фауны (более 10% семейств и до 30% родов) имело место на рубеже силура и девона, однако к середине девона ее таксономическое разнообразие вновь несколько возросло. В начале позднего девона оно опять начинает снижаться и особенно резко падает на рубеже франского и фаменского веков, когда вымирает более 15% семейств, более 40% родов и до 65% видов морских животных. Биотический кризис на рубеже девона и карбона унес более 15% семейств и более 30% родов морской фаменской фауны.
В течение каменноугольного и большей части пермского периодов уровень таксономического разнообразия морской биоты существенно не изменился (эпоха "стазиса"), но в конце перми произошло величайшее вымирание организмов, унесшее на протяжении нескольких миллионов лет около 40-50% семейств, более 70% родов и 95% видов морской фауны.
В первой половине триаса таксономическое разнообразие биоты вновь несколько возросло, достигнув 2/3 от наивысшего пермского уровня, но в конце триаса разразилось новое великое вымирание и исчезло около 20% семейств и около 50% родов морских организмов. Затем на протяжении юры и мела происходил быстрый и почти неуклонный (если не считать небольшого кризиса на рубеже юры и мела) рост таксономического разнообразия биоты. В конце мела - в середине маастрихтского века началось и на рубеже мела и палеогена (65 млн. лет назад) завершилось последнее великое вымирание, в результате которого разнообразие животного мира сократилось на уровне семейств на 16-17%, родов - почти на 50% и видов - на 77% и полностью исчезли динозавры, аммониты, белемниты, иноцерамы, рудисты и пр. После этого вымирания возобновился продолжающийся поныне значительный рост числа таксонов, несколько осложнявшийся относительно малым биотическим кризисом на рубеже эоцена и олигоцена, и к концу кайнозоя количество семейств, родов и видов намного превысило эти показатели для середины Маастрихта.
Обсуждению возможных причин великих вымираний посвящена огромная литература, и было высказано множество различных гипотез. А.С.Алексеев (1998) предлагает различать среди них гипотезы, связывающие феномен вымираний с внешними (по отношению к Земле) и внутренними причинами. К первой группе относятся галактические гипотезы, связывающие массовые вымирания с возможными изменениями потока космических лучей, интенсивности магнитного и гравитационного полей, пересечением Солнечной системой в течение галактического года газопылевых струй Галактики; гипотезы, предполагающие связь эпох массовых вымираний с влиянием колебаний солнечной активности, напряженности магнитного поля Солнца, эффективностью озонового экрана, ультрафиолетового излучения Солнца и пр. Импактные гипотезы объясняют великие вымирания падениями на Землю одного или нескольких крупных астероидов или комет, вызыварших так называемую "иридиевую аномалию", проявления метаморфизма высокого давления и пр.
Ко второй группе относятся гипотезы, предполагающие связь великих вымираний с фазами диастрофизма, пароксизмами вулканических извержений, фазами резкого повышения радиационного фона, усилением спрединга и рифтинга и вызванного временным усилением спрединга выносом из рифтовых зон океанов и континентов больших количеств радиоактивных элементов или токсических веществ, с инверсиями геомагнитного поля Земли и изменениями их частоты, с эв-статическими колебаниями уровня океана, с неблагоприятными временными резкими колебаниями состава гидросферы и атмосферы, а также с влиянием таких биотических факторов, как эволюционный дисбаланс сообществ организмов, исчерпание пищевых ресурсов и пр.
В настоящее время проблема причин изменения таксономического разнообразия биот, в том числе великих вымираний, еще далека от разрешения. Так, например, весьма модна в последнее время гипотеза Л.Альвареса о связи мел-палеогенового вымирания с одним или несколькими од-
новременными катастрофическими импактными событиями, происшедшими на этом рубеже. Эта гипотеза опирается на эмпирически установленные факты (иридиевая аномалия, признаки шокового метаморфизма, обнаружение нескольких взрывных кольцевых структур, трактуемых в качестве астроблем) и дает, казалось бы, убедительное подтверждение космической причины этого великого вымирания. Однако, во-первых, предполагаемое импактное событие совпало во времени лишь с самым концом эпохи вымирания, продолжавшейся несколько миллионов лет; во-вторых, эта гипотеза оказывается неприменимой к ряду других великих вымираний в фанерозое, в том числе - к величайшему среди них вымиранию на границе перми и триаса, так как иридиевой аномалии и признаков шокового метаморфизма в отложениях, отвечающих этим рубежам, не установлено. Кроме того, следует отметить, что космическая природа всех крупных взрывных кольцевых структур с признаками шокового метаморфизма оспаривается некоторыми авторитетными исследователями (П.Н.Кропоткин, А.А.Маракушев и др.).
Е.Е.Милановский (1999) считает, что многие факторы, предполагаемые разными гипотезами о внутренних (т.е. "земных") причинах вымираний, такие, как фазы диастрофизма (расширения или сжатия земной коры), вулканические пароксизмы, эвстатические колебания уровня Мирового океана, изменения частоты геомагнитных инверсий, глобальные изменения климатической обстановки на Земле, находятся в тесной взаимосвязи и в конечном счете порождаются общепланетарными тектоническими процессами, прежде всего, глобальной геотектонической цикличностью различных порядков и продолжительности.
Е.Е.Милановский сопоставляет крупнейшие циклы геотектонического (и в более широком плане - геологического) развития Земли в фанерозое с крупнейшими глобальными циклами изменений таксономического разнообразия биоты, которые, как правило, развиваются по следующему "сценарию" (Алексеев, 1998): 1) Период низкого таксономического разнообразия биоты (от одного до нескольких миллионов лет). 2) Период быстрого роста ее таксономического разнообразия (диверсификации). 3) Продолжительный или короткий период "стазиса", во время которого СОХра-
няется достигнутый высокий уровень таксономического разнообразия. 4) Период биотического кризиса, во время которого происходит либо постепенное, либо многофазное значительное снижение таксономического разнообразия биоты (продолжительностью от первых миллионов лет до 10-15 млн. лет), приводящее в итоге к великому вымиранию.
Среди великих вымираний фанерозоя выделяются два величайших: 1) на рубеже венда и кембрия (570 млн. лет назад), когда исчезла практически вся бесскелетная фауна венда, а затем в течение не более 20-25 млн. лет происходила быстрая диверсификация морской биоты, достигшей наибольшего разнообразия в ботомском веке; 2) в конце поздней перми - на рубеже перми и триаса (250 млн. лет назад), когда вымерла почти половина семейств, почти три четверти родов и до 95% видов морской фауны. Оба этих величайших вымирания совпадали с самыми важными рубежами в геотектоническом (в целом и геологическом) развитии Земли в фанерозое,- соответственно с границами: 1) между позднерифейско-вендским (байкальским) и палеозойским мегациклами; 2) между палеозойским (герцинским) и мезозойским мегациклами. Немного уступают этим главным биотическим катастрофам по своей роли в истории органического мира Земли несколько эпох великих вымираний, во время которых исчезала почти половина (не менее 40%) всех родов и двух третей всех видов ископаемой морской фауны. К ним можно отнести: 1) несколько тесно сближенных во времени великих вымираний в середине и конце кембрия; 2) великое вымирание на рубеже ордовика и силура; 3) тесно сближенные великие вымирания в конце девона (на рубежах франского и фаменского, фаменского и турнейского веков); 4) великое вымирание в конце триаса и, наконец, 5) великое вымирание в конце мела. Все эти события совпадают с границами циклов Бертрана: 1) с завершением салаирского цикла и его границей с каледонским; 2) и 3) с рубежами между каледонским и герцинским циклами в тех регионах подвижных поясов, где каледонский цикл завершился сравнительно рано таконской фазой складчатости (например, в Казахе-
ком нагорье и Северном Тянь-Шане), либо, напротив, - наиболее поздно (в позднем девоне, как например, в эпоху акадской орогении в Аппалачах); 4) с рубежом между ранне- и позднекимме-рийским циклами (индосинийская орогения в конце триаса, в Индокитае, на Таймыре, Новой Земле, Мангышлаке и пр.) и, наконец, 5) с рубежом между позднекиммерийским и альпийским циклами (с эпохой ларамийской складчатости в Кордильерской системе и других регионах Тихоокеанского и Средиземноморского поясов). Как правило, каждая из эпох великих вымираний совпадала во времени с эпохами сжатия земной коры (фазой или несколькими сближенными фазами складчатости), завершающими геотектонический цикл Бертрана в том или ином подвижном поясе или его крупном регионе, а также с совпадающим с ним во времени тектоноэвстатическим понижением уровня океана. Самый конец эпох великих вымираний отличался сменой падения этого уровня началом его последующего подъема, а в некоторых случаях - началом короткой, но грандиозной по своему масштабу фазы базальтовых излияний в консолидированных областях континентов (например, в самом начале раннего триаса в Сибири и в самом начале палеоцена в Индос-
; тане и пр.). Это указывало на переход от эпохи или фазы преобладающего глобального сжатия земной коры к последующей эпохе или фазе ее преобладающего расширения.
Приведенные данные, на взгляд Е.Е.Милановского, достаточно убедительно свидетельствуют
. о приуроченности этих эпох к завершающим, контракционным фазам крупных геотектонических циклов. Однако он не исключает возможности связи этапов эволюции биоты, и в частности эпох вымираний, с космическими событиями, происходившими на Солнце, в Солнечной системе и Галактике. По-видимому, космические события в значительной мере контролировали ход многих геологических процессов, происходивших на Земле, а также их периодичность.
Палеонтологический музей Томского государственного университета, как и другие подобные музеи, является лучшим местом для показа последовательных изменений ископаемых и, в целом, развития органического мира в геологическом прошлом. С помощью остатков фауны и
флоры, ДОбыТЫХ ИЗ обнажений И глубин Земли (керновый материал), воссоздается уникальная си-
стема координат, дающая возможность сопоставить в пространстве и времени геологические формации, к которым приурочены не только указанные органические остатки, но и различные полезные ископаемые. Бесценные свидетельства развития биосферы нашей планеты прекрасно представлены в экспозициях Палеонтологического музея ТГУ, по праву считающимся одним из фундаментальных музеев не только в России, но и во всем мире.
Основателем и первым научным руководителем музея, открытого в 1926 г., был профессор В.А.Хахлов, который возглавил впоследствии созданную им школу палеоботаников. В последнее десятилетие музей значительно обновлен и преобразован. Он выполняет различные взаимосвязанные функции: популяризаторскую (демонстрация палеонтологического материала для представления об эволюции биосферы), научную (углубленное изучение ряда групп ископаемой фауны и флоры), а также просветительскую, являясь частью образовательного комплекса Томского государственного университета.
В витринах и диорамах представлены остатки руководящих ископаемых растений и животных фанерозоя, смоделированные художниками ландшафты прошлых геологических эпох. Весьма информативны и зрелищны четыре диорамы с реконструкциями основных периодов истории Земли, а также витрины, украшенные картинами растительных ландшафтов и отпечатками растений, собранных в разных регионах Сибири. Они отражают четыре этапа развития высших растений в фанерозое (позднесилурийский-раннедевонский - "псилофиты" или риниофиты; позднедевоне*-кий-раннепермский - плауновидные, хвощевидные, папоротниковидные; позднепермский-ранне-меловой - голосеменные; позднемеловой-современный - покрытосеменные растения). В музее создана витрина, на которой показана эволюция растений с распределением их по биогеографическим провинциям. Здесь же приведены схемы строения растений, сопровождаемые замечательными по сохранности их отпечатками (рис. 77, цв. вкл.).
Сибирь богата многочисленными палеонтологическими остатками, основные местонахождения которых представлены в отдельной витрине. Здесь (снизу вверх) на отдельных полках показаны различные остатки фауны и флоры: от девона до неогена включительно, обнаруженные преимущественно на юге Западной Сибири. Раннедевонская флора в основном состоит из "псилофи-тов" и водорослей лагунных отложений Минусинских впадин. Отпечатки рыб найдены в породах изыкчульского горизонта Хакасии. Брахиоподы, мшанки и криноидеи, выставленные в этой витрине, собраны из отложений опорного разреза нижнего карбона Лагерного сада Томска. Большой интерес вызывают находки юрских растений, представленные экспонатами из обнажений Кузбасса и в керновом материале из разрезов многих скважин Томской области. Хорошо сохранившиеся отпечатки растений неогена, собранные из разных местонахождений преимущественно Томской области, завершают верхнюю часть этой экспозиции. Самыми древними экспонатами музея являются остатки жизнедеятельности древних "сине-зеленых водорослей" - цианобионтов, возраст которых исчисляется более 600 млн. лет ("невландиевая" проблематика, строматолиты).
Коллекции музея интенсивно пополняются. Ежегодные полевые экскурсии сотрудников Сибирского палеонтологического научного центра (СПНЦ) совместно со студентами добавляют .$ музей новые экспонаты. Особенно важны последние находки на юго-востоке Западной Сибири остатков рептилий и млекопитающих. Летом 1995 г. экспедицией СПНЦ были обнаружены в окрестностях с. Шестаково по р. Кие (левый приток р. Чулыма) два новых местонахождения ран-немеловых позвоночных. В последующие годы благодаря тщательным и квалифицированным раскопкам найдены остатки различных животных, большая часть которых принадлежит раннемело-вым динозаврам - пситтакозаврам. Летом 1999 г. сотрудникам СПНЦ удалось найти два целых
скелета пситгакозавров, примерно до 2 м в длину, 1,5 М В ВЫСОТу (рИС. 78). НаХОДКИ ЭТИХ реПТИЛИЙ
ИЗВеСТНЫ ранее ИЗ НИЖНемеловых отложений на северо-западе Китая и в Монголии. По-видимому, в раннемеловое время существовал единый ареал их обитания от Сибири до Китая.
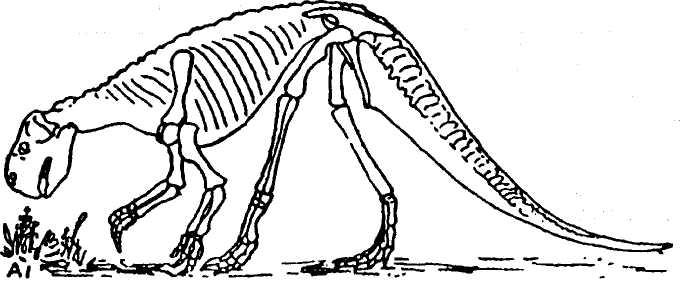
Рис. 78. Реконструкция скелета мелового травоядного динозавра рода Psittacosaurus (no W.Gregory, 1927)
Помимо находок раннемеловых динозавров, в четвертичных отложениях в ряде местонахождений юго-востока Западной Сибири обнаружены остатки мамонтовой фауны, которые дали возможность изучить эту группу млекопитающих и сконструировать в музее их скелеты. Наиболее интересны реконструкции скелетов мамонта, бизона и шерстистого носорога - представителей хоботных и копытных позднего плейстоцена юго-востока Западной Сибири (рис. 79, цв. вкл.).
Посещение демонстрационного зала Палеонтологического музея с его красочно оформленными диорамами, витринами, макетами, скелетами древних животных - это увлекательнейшее путешествие в прошлое Земли. Оно способствует осмыслению уже имеющихся знаний, расширению горизонтов миропонимания, повышает интерес к познанию развития органического мира нашей планеты.