

Органи чуття
Органи чуття по-різному розвинені в окремих рядах ссавців. На перше місце для жителів відкритих просторів виходить зір, для нічних і присмеркових тварин – жителів лісових і чагарникових біотопів, норників і жителів водойм – нюх і слух.
Нюх ссавців ефективніший, ніж у інших наземних хребетних. Велика роздільна здатність хеморецепторів дозволяє розрізняти окремі специфічні речовини (запахи) або їх поєднання, характерні для виду, групи особин або індивідів. У різних рядах і в окремих видів ссавців гострота нюху неоднакова. Сумчасті, комахоїдні, гризуни, неповнозубі, більшість хижаків і копитних – так звані макросматики, відрізняються високо розвиненим нюхом; він використовується при орієнтації в просторі, пошуках їжі, у міжвидових і внутрішньовидових зв’язках. Більшість приматів і ряд інших ссавців володіють менш чутливим нюхом (мікросматики).
Органи нюху розташовуються у верхнє-задній частині носової порожнини, де виникає складна система раковин, вкрита слизовою оболонкою з нюхового епітелію з рецепторними клітинами з характерними волосками. Аксони цих клітин об’єднуються в групи, утворюючи волокна, що входять в нюхові цибулини. Останні через ланцюг нейронів з’єднуються з центрами головного мозку. Складність будови нюхових раковин відповідає гостроті нюху (рис. 27).
У
Рис. 27. Поперечний розріз задньої частини носової порожнини ссавців.
А – мікросматик (людина); Б – макросматик (козуля)
китоподібних наявність нюху і смаку заперечувалась і їх називали аносматиками. Недавні дослідження довели, що дельфіни мають пахучі залози, що відкриваються біля анального отвору; тварини здатні визначати за слідами їхнього секрету напрямок табуна, що пройшов; вони сприймають запах крові як сигнал небезпеки. У ротовій порожнині вусатих китів є парні заглибини на кінці верхньої щелепи, гомологічні якобсонову органу інших хребетних. У основі язика зубатих китів розташовані довгасті ямки, що нагадують смакові сосочки інших ссавців. Очевидно, з їх допомогою кити розпізнають запахи і орієнтуються , розрізняючи течії з різним хімізмом. Мозок китоподібних, хоч і відрізняється редукцією нюхових долей, але зберігає в корі півкуль структури, пов’язані з аналізом хімічних сигналів.Слух в житті ссавців відіграє важливу роль. Цьому відповідає і складне влаштування голосового апарату, здатного видавати різноманітні звуки, часто утворювати складні поєднання, організовані в часі. За шириною звукового діапазону ссавці переважають птахів, широко використовуючи як ультразвуки (вище 20 кГц), так і низькі частоти. Слух і звукова сигналізація обслуговують найважливіші життєві явища – пошуки їжі, виявлення небезпеки, розпізнавання свого і чужого видів, різницю індивідів в групі (стаді або зграї), відносини батьків і малят й багато іншого. Особливості слуху відрізняються у різних рядах. Так, для ехолокації кажани використовують переважно ультразвукові частоти в межах 40―80 кГц, але видають і низькочастотні звуки до 12 Гц (не чутні нашим вухом інфразвуки). Ще ширшим є звуковий діапазон вусатих китів – від декількох герц до двохсот кілогерц. Вусаті кити видають звуки низької частоти (1―2 кГц) значної сили і тривалості. Здатністю до ехолокації наділені комахоїдні (землерийки) і деякі гризуни, що ведуть норний спосіб життя. Різні діапазони використовуються одним видом для різних цілей – ехолокація і пошук здобичі на високих і ультразвукових частотах, спілкування з особинами свого виду – на відносно низьких.
Орган слуху ссавців складається з трьох відділів: зовнішнього, середнього і внутрішнього вуха. Зовнішнє вухо (вушна раковина) і зовнішній слуховий прохід своєрідна звукова антена – фільтр, здатну вбирати і посилювати біологічно важливі для виду звуки, послаблюючи сторонні шуми. Цю ж роль виконує середнє вухо (рис. 28), в якому три слухові кісточки: молоточок, ковадло і стремінце – утворюють систему важелів, яка передає коливання більшій за величиною барабанній перетинці на меншу за площею мембрану овального віконця внутрішнього вуха, посилюючи ці коливання.
К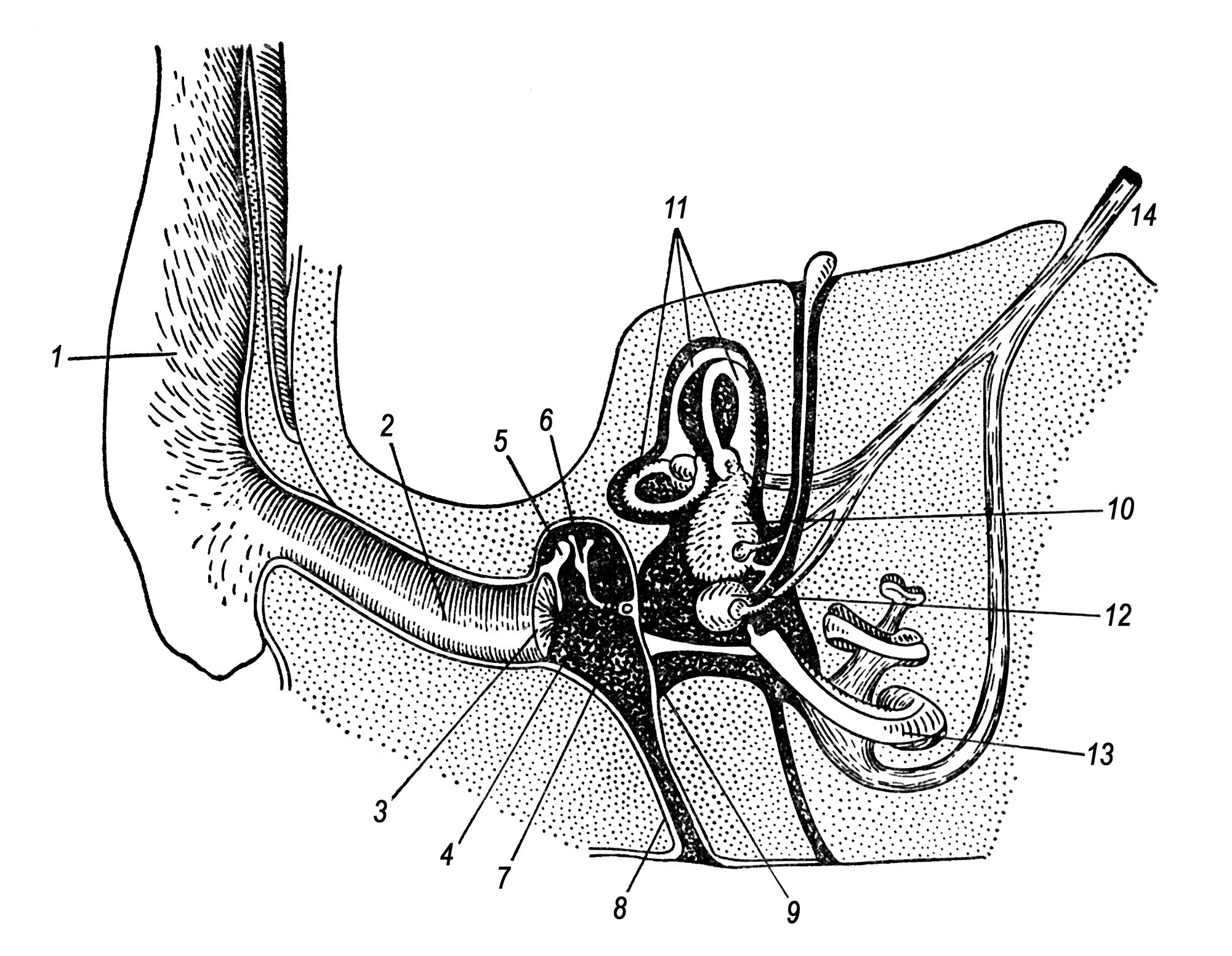 істковий
барабан, в якому знаходиться порожнина
середнього вуха, утворює ряд камер –
резонаторів, що посилюють біологічно
важливі звуки. У деяких видів, особливо
норників, вони іноді заповнені губчатою
кістковою масою, що гасить сторонні
шуми. Середнє вухо євстахієвою трубою
з’єднується з задньою частиною ротової
порожнини, що забезпечує вирівнювання
тиску повітря по обидва боки барабанної
перетинки.
істковий
барабан, в якому знаходиться порожнина
середнього вуха, утворює ряд камер –
резонаторів, що посилюють біологічно
важливі звуки. У деяких видів, особливо
норників, вони іноді заповнені губчатою
кістковою масою, що гасить сторонні
шуми. Середнє вухо євстахієвою трубою
з’єднується з задньою частиною ротової
порожнини, що забезпечує вирівнювання
тиску повітря по обидва боки барабанної
перетинки.
В
Рис. 28. Схема органу слуху ссавця:
1 – зовнішнє вухо; 2 – зовнішній слуховий прохід; 3 – барабанна перетинка; 4 – порожнина середнього вуха; 5 – молоточок; 6 – ковадло; 7 – стремінце; 8 – євстахієва труба; 9 – кругле вікно; 10 – овальний мішечок; 11 – півколові канали; 12 – круглий мішечок; 13 – завитка; 14 – слуховий нерв.
нутрішнє вухо розташоване в товщі скроневої кістки (в її кам’янистій частині) і складається з вестибулярного і слухового відділів. Вестибулярний відділ включає три півколові канали і овальний мішечок; він є органом рівноваги і сприйняття просторового положення тіла. Слуховий відділ утворений круглим мішечком і пов’язаною з ним завиткою, в якій розташований кортіїв орган; функції останнього полягають в первинному аналізі, переважно частотному, і кодуванні звукових сигналів, які після обробки передаються в слуховий центр (аналізатор) мозку. Завитка – спірально вигнута перетинчаста трубка, яка лежить в кістковому футлярі, – заповнена ендолімфою. В центрі завитки по всій її довжині йде базальна мембрана, на якій впоперек натягнуті фібрили (слухові струни). До них дотикаються чутливі клітини кортієвого органу, що сприймають коливання слухових струн, настроєних на різну частоту. Імпульси, сприйняті чутливими клітинами, передаються нейронам, аксони яких утворюють слуховий нерв. Такий механізм забезпечує тонкий аналіз частотного спектра і часової організації звукового сигналу, що сприйнятий зовнішнім вухом і переданий посиленим через середнє внутрішньому вуху.Коливання голосових зв’язок верхньої гортані приводить до утворення звуків у ссавців. Ультразвукові сигнали кажанів генеруються апаратом рота або носа. У китоподібних в утворенні звуків беруть участь гортань в цілому, краї черпакуватих хрящів, повітряні мішки носового проходу і зовнішнє дихало. Окрім голосу, деякі ссавці використовують механічні звуки: клацання і скрегіт зубами (хижаки, деякі гризуни, копитні, примати), стукіт рогами, удари ногами об землю (багато норних тварин, копитні), шум від тертя голок (дикобраз) і т. п.
Зоровий аналізатор є третім основним органом чуття ссавців. Для деяких звірів, що ведуть переважно денний спосіб життя і живуть у відкритих біотопах, більша частина навколишньої інформації поступає через зоровий канал. Значення зору зменшується у жителів лісів, заростей і трав’яного покриву. У норників очі деколи перестають функціонувати , заростаючи шкірою (деякі кроти, сліпаки), або реєструють лише зміну освітлення (сліпушок, прометеєва полівка). У китоподібних очі використовуються лише для ближньої орієнтації. Очі ссавців розташовані або по боках голови, забезпечуючи майже кругове поле зору, при якому бінокулярний зір обмежений невеликим сектором, або фронтально. У останньому випадку загальний огляд скорочується, але поле бінокулярного зору збільшується. Перший тип переважає у копитних і гризунів, що постійно очікують нападу ворогів; другий характерний для мавп, що ведуть деревний спосіб життя, яким необхідно точно визначати відстань при стрибках з гілки на гілку, і для частини хижаків, особливо котячих, які, нападаючи з засади, мають точно фіксувати відстань до жертви. Відносна величина очей зростає у тварин з більш гострим зором і у тварин з нічною активністю.
Око ссавців має зовнішню оболонку (склеру) з волокнистої тканини. В передній частині склера переходить в прозору роговицю. Під склерою лежить судинна оболонка з кровоносними судинами, що постачають необхідні речовини. Між склерою і судинною оболонкою у деяких тварин є шар клітин з кришталиками, що утворює відбиваюче промені дзеркальце (tapetum), яке обумовлює „світіння” ока відбитим світлом (хижі, копитні). Потовщуючись, судинна оболонка спереду переходить в райдужну і війчасте тіло (м’язи), за допомогою якого відбувається акомодація ока зміною форми кришталика. Райдужна оболонка відіграє роль діафрагми, регулюючи освітленість сітківки зміною величини зіниці. Кришталик лінзоподібної форми відносно малий у денних ссавців і різко збільшується у тварин з нічним способом життя. До внутрішньої сторони судинної оболонки прилягає сітківка, що складається з зовнішнього пігментного і внутрішнього світлочутливого шарів. Колбочки не мають жирових крапель. Відмінності між видами зводяться до варіацій у співвідношенні паличок і колбочок, коливаннях загального числа рецепторних клітин і їх кількості на одне волокно зорового нерва. У норних тварин кількість рецепторних клітин і волокон нерва мінімальна: у сліпака у всій сітківці 800 тис. рецепторів і 1900 волокон в зоровому нерві (співвідношення 420:1). У нічних видів і жителів заростей воно вище: у їжака 6,7 млн. рецепторів на 8400 волокон (760:1), у жовтогорлої миші 19,6 млн. і 28 800 (680:1). Ще більше це число у жителів відкритих ландшафтів: так, у зайця-русака 192,6 млн. рецепторів і 167 400 волокон (115:1). У макаки-резуса (примати) 124,4 млн. рецепторів на 1,2 млн. волокон (105:1), а у кажана (рукокрилі) лише 8,9 млн. рецепторів на 6900 волокон (1130:1). Кількість рецепторних клітин, що в середньому припадають на одне нервове волокно зорового нерва, найменше у приматів; це дозволяє виявити більше деталей в об’єктах.
Багато ссавців володіють здатністю розрізняти кольори, але, очевидно, слабше, ніж птахи. З цим пов’язано в середньому менш різноманітне забарвлення ссавців. В той же час ссавці розпізнають особливості форми предметів або їх частин, а також рухи, позу і міміку. Це забезпечено не ускладненням будови сітківки, а зоровим аналізатором в головному мозку, який у ссавців складніший, ніж у інших хребетних. Основну роль відіграє зоровий центр кори півкуль переднього мозку, тоді як значення зорової кори середнього мозку (передніх горбів чотиригорбикового тіла) зменшується. Перенесення основної обробки зорової інформації у центр півкуль переднього мозку відкрило можливості не тільки для візуальної орієнтації в просторі, а і для ускладнення і збагачення зорових зв’язків між особинами. Виникли і широко використовуються ссавцями „мова форми, поз, жестів і міміки”. Це відіграє роль у врегулюванні відносин у популяціях і утворенні угруповань з узгодженою поведінкою (див. нижче). Забарвлення і форма тіла тварин під впливом природного добору набули маскуючого значення (критичне забарвлення) або були демонстрацією при погрожувальній поведінці.
На ділянках тіла, що часто стикаються з навколишніми предметами, ссавці мають особливо довгі і жорсткі волосини – вібриси. Їх корені пов’язані з закінченням нервів. Особливого розвитку вібриси досягають звичайно на морді.
Поведінка і спосіб життя
Загальні особливості поведінки. Високоорганізовані нервова система і органи чуття забезпечують складне відображення зовнішнього світу в мозку і відповідно гнучку поведінку більшості ссавців. У її основі лежать механізми, властиві всім тваринам: „біологічний годинник”, що керує циркадними ритмами, сукупність простих безумовних рефлексів і їх складних поєднань – інстинктів – і, накінець, здатність до тонкого аналізу і сприйняття зовнішніх впливів, що супроводжуються утворенням умовних рефлексів (тимчасових зв’язків) і накопиченням індивідуального досвіду. Це неможливо без існування спадкової, генетично закріпленої пам’яті (основи безумовних рефлексів і інстинктів) і біжучої пам’яті у вигляді тимчасових зв’язків. Те і інше у сукупності становить „оперативну пам’ять”. На її основі оцінюється ситуація за сприйнятою органами чуття інформацією про стан середовища і організма і вибирається оптимальний варіант дії.
Ссавці вирізняються серед інших тварин багатством оперативної пам’яті і відповідно складною поведінкою. Її пристосувальний характер посилюється так званим імпринтингом, або запам’ятовуванням – стійким, звичайно збереженим до кінця життя впливом перших, сприйнятих в дитячому віці, „вражень” на всю наступну поведінку. Імпринтинг- це добудова і пристосування функціональних систем організму щодо конкретних умов вибраного місцеперебування. Тому нервова діяльність ссавців відрізняється високим ступенем рухливості, багатством і складністю зв’язків з оточуючим середовищем і одночасно високим рівнем пристосування до місцевих умов. Утворюючи групи індивідів, ссавці використовують ресурси середовища і зменшують несприятливі впливи, колективно створюючи запаси їжі, влаштовуючи нори і інші прихистки зі сприятливим мікрокліматом, об’єднуючи зусилля в захисті від ворогів і пошуках їжі. Такі зародки „соціальних” елементів в популяціях ссавців представлені більш різноманітно у порівнянні з плазунами і птахами.
Вроджена, або інстинктивна, діяльність ссавців складніша і рухливіша, ніж у інших хребетних. Виникає і широко використовується здатність передбачати хід багатьох подій, що повторюються, і приймати відповідні рішення (екстраполяційні рефлекси). Вона, очевидно, відсутня у інших хребетних, властива різним групам птахів і значно багатше і чіткіше виражена у ссавців.
Популяційна організація і спосіб життя. Ускладнення внутрішньовидових зв’язків супроводжується утворенням тимчасових або стійких угруповань особин на основі спілкування та узгодженості в поведінці. Так виникає „етологічна” структура популяцій, що відповідає способу життя і способу розмноження виду. Ссавці, що живуть в норах або користуються постійними сховищами, звичайно ведуть поодинокий або сімейний спосіб життя, займають певні ділянки, що захищаються від проникнення чужинців (багато гризунів, комахоїдні, хижаки). Перевага такого „землекористування” полягає у послабленні конкуренції за їжу і сховища, і у відносно рівномірному використанні території. Великі за розміром рухливі тварини, особливі копитні, утворюють кочуючі стада або зграї. Кочівлі зменшують небезпеку виснаження пасовиськ, а об’єднання у групи збільшує захист проти хижаків. У китоподібних і ластоногих об’єднання у групи збільшує можливість виявлення скупчень корму.
Проміжне положення між поодиноким і стадним способом життя займають колоніальні поселення гризунів (бабаків, ховрахів і ін.) або зайцеподібних (пищух). Вони існують у місцях з достатньою кількістю корму і забезпечують кращі можливості для виявлення хижаків, особливо птахів. В умовах тісного співжиття і постійного спілкування сусідів ці тварини в межах поселення мають свої індивідуальні або сімейні ділянки. Часто тут поселяються і інші види, що ведуть поодинокий спосіб життя, і яких приваблюють сприятливі умови (їжа, сховища) таких колоній.
У групах тварин з номадним (кочівним) способом життя і у поселеннях видів, що ведуть поодинокий або сімейний спосіб життя, утворюються групи особин, які підтримують більш часті контакти і нерідко діють сумісно. Такі „парцелярні угруповання”, „великі сім’ї”, або клани, ефективніше забезпечують розмноження, одночасно впорядковуючи використання кормів, полегшуючи створення сховищ, прихистків і стежок в місцях перебування. Контакти і часті взаємодії тварин у таких угрупованнях на більш широкій основі забезпечують виховання молоді і збагачення її досвідом попередніх поколінь. Структура популяцій виникає у ході спілкування тварин, відображаючи їх видові і індивідуальні особливості. Основою утворення популяційної структури служить „ранжування” особин, яке відображає різний стан особин в групі: „домінант”, „субдомінінти” і „підлеглі”. Система „рангів” в популяції звичайно складна і завжди рухома. У ссавців, особливо стадних, вона набуває особливої складності. Зустрічаються „колові” системи супідрядності і існують паралельні, не співпадаючі системи супідрядності, що обслуговують різні сторони життя групи (рис. 29).

Прямолінійне домінування
Складне домінування
Рис. 29. Системи „ранжування” (супідрядності) в різних популяційних угрупованнях
Буквами позначені окремі особини, цифрами – кількість випадків домінування.
Кільцеве домінування
Набуте положення в групі звичайно пов’язане зі спадковими особливостями, станом особини та її досвідом. Домінант втрачає ранг в результаті помилки, хвороби або появи більш сильного суперника. Така рухомість системи рангів забезпечує високі здатності групи до пристосувань в змінених умовах існування. Ранжування звичайно супроводжується сутичками між претендентами, особливо гострими у стадних тварин, які в період розмноження утворюють гареми. Емоційно напружені поєдинки самців часто в цей час могли б бути смертельними для суперників, якби у ході природного добору знаряддя нападу не набули ”турнірного”, відносно безпечного характеру, або не виникли захисні пристосування, подібні „калкану” кабанів – потужної, пружної сполучнотканинної жирової подушки на плечах, що захищає від ікол суперника.
В деяких групах спостерігається розподіл обов’язків. Так, у зграях (прайдах) левів самці переважно охороняють мисливські території від вторгнення чужинців (членів сусідніх прайдів), а самки здобувають їжу і турбуються про молодь. У зграях (великих сім’ях) вовків при нападі на велику здобич одні заганяють її, тоді як інші прагнуть перерізати їй шлях або нападають із засідки.
Складна внутрішньопопуляційна структура змінює характер дії природного добору: поряд з індивідуальним відбувається і добір груп з кращою організацією. Поділена на парцелярні угруповання популяція не втрачає своєї цілісності, яка забезпечується загальною системою внутрішньопопуляційних зв’язків (хімічних, візуальних, акустичних), влаштуванням спільних нір, стежок, гнізд і т. п. На зайнятій популяцією території виникає „сигнальне поле”, зв’язуюче парцели в єдину популяцію (Наумов, 1975). Вивчення системи сигналізації і сигнальних полів має практичний інтерес, відкриваючи можливості управління розміщенням, чисельністю і поведінкою корисних і шкідливих диких тварин з допомогою технічного відтворення сигналів або їх систем. Існує обмін особинами між популяціями; він здійснюється при розселенні молоді. При дозріванні змінюється стан молодих тварин (підвищується рівень і мінливість їх метаболізму, зростає нервова збудливість). В цей час вони покидають знайому територію в пошуках партнера і нових місць перебування. Як показало мічення, при розселенні молодняк здійснює звичайно ненаправлені переміщення, які в десятки і сотні разів перевищують рухливість дорослих.
Біологічні (річні) цикли неоднакові у ссавців з різним способом життя. Час народження молоді завжди припадає на найбільш сприятливий для їх росту і розвитку період, у більшості випадків на весну - початок літа. Час парування обумовлений тривалістю вагітності (інколи збільшується діапаузою) і може припадати на різні сезони року: весною (у гризунів і комахоїдних), взимку (у собачих), восени (у копитних), влітку (у куниць) або відразу після родів (ластоногі). У ссавців з декількома приплодами на рік (дрібні гризуни) розмноження за сприятливих умов може тривати протягом всього року (миші, полівки, лемінги).
Після підростання молоді у тварин з поодиноким способом життя сім’ї розпадаються і відбувається розселення (дисперсія) молоді. У стадних форм молодь або продовжує триматися разом з самками, або утворює окремі групи. Восени ссавці готуються до зимівлі; в цей час тварини накопичують жирові резерви, змінюють літній волосяний покрив на зимовий, деякі види роблять запаси корму на зимовий період. Одночасно відбувається перебудова популяційної структури; до зимового періоду дрібні групи тварин часто об’єднуються в більші за розміром групи. У норах гризунів (мишей, полівок), які влітку живуть поодинокими сім’ями, знаходили по 20-30 тварин в одному прихистку. Багато копитних до зими об’єднуються у великі табуни, що сприяє міграціям і перебуванню в місцях годування. Утворення груп дозволяє хижакам (вовкам і ін.) добувати великих за розмірами тварин. Лише дрібні хижі (кунячі), активні в зимовий період комахоїдні, деякі гризуни і зайці продовжують поодиноке існування.
У сплячку або зимове заціпеніння впадає багато ссавців, але характер цих явищ в різних рядах неоднаковий. Сплячка відсутня у копитних, ластоногих і китоподібних, для яких властиві кочівлі (номадний спосіб життя). У білок в суворі похолодання під час зими лише зменшується активність. У деяких хижаків (ведмеді, борсуки і ін.) сплячка проявляється у пригніченні рухової активності, при цьому зберігається високий рівень обміну речовин і температури. Більш глибокий сон властивий деяким гризунам (хом’якам) і кажанам. Справжня сплячка з повним заціпенінням і різким зниженням температури тіла властива деяким комахоїдним, наземним білячим (бабакам і ховрахам), соням і тушканчикам. Вона властива не лише для зимового періоду, а й літньої посухи.
Міграції властиві широкому колу ссавців. Хижаки в пошуках здобичі змінюють місця перебування в різні сезони, переміщуються на порівняно невеликі відстані. Обмежені сезонні кочівлі здійснює багато гризунів. Значні кочівлі властиві більшості стадних видів. На сотні і навіть тисячі кілометрів мігрують деякі копитні (північний олень), ластоногі і китоподібні, а також деякі кажани. Сезонні кочівлі, як і перельоти птахів, повторюються щорічно і здійснюються певними міграційними шляхами. Сезонні міграції потрібно відрізняти від розселення молоді, які в роки підйомів чисельності часто набувають характеру масового виселення (еміграція). Саме такі переважно „міграції” білок і нашестя лемінгів; песці протягом таких „сезонних кочівель” проникають в південну тайгу.
Географічне поширення. Вивчення особливостей поширення ссавців, як і інших груп тварин, дозволило провести зоогеографічне районування Землі. Для Австралійської області характерно панування різноманітних сумчастих, тільки тут збереглися найпримітивніші ссавці - однопрохідні. З плацентарних тут найбільш різноманітні кажани. Небагаточисельні гризуни об’єднуються в ендемічну родину Hydromyidae. Людина завезла сюди щурів і собаку динго, тут були акліматизовані кролик, багато копитних, деякі хижі та ін.
Неотропічна область (Центральна і Південна Америка) має багату і різноманітну фауну ссавців. Близько 30 видів сумчастих опосумів екологічно заміщають практично відсутніх комахоїдних. Тільки тут зустрічаються мурашкоїди, лінивці, броненосці (ряд неповнозубі). Багато рукокрилих (включно з кровосисними формами – вампірами). Ендемічні різноманітні гризуни (агуті, свинки, шиншили, нутрії, деревні дикобрази і ін.) і широконосі мавпи (капуцини, ігрунки і ін.). З непарнокопитних зустрічаються лише тапіри. Малочисельні і парнокопитні: своєрідні свинячі – пекарі, безгорбі верблюди – лами, маленькі за розмірами олені Pudu.
У Ефіопській області (Африка, крім північних її районів) ендемічні дамани і трубкозубі. Різноманітні комахоїдні (златокроти, видрові землерийки, стрибунцеві та ін.) і гризуни (шипохвості летяги, земляні білки, кафрський довгоніг, землекопи та ін.). Парнокопитні представлені значною кількістю різноманітних антилоп, буйволом, жирафами, бегемотами, свинями; непарнокопитні – носорогами і зебрами; хоботні – слонами. Різноманітні хижі (особливо вівери), але частина з них виходить за межі області. (лев, леопард, гепард, гієни). Багато мавп родини мартишкові; ендемічні дуже рідкісні у теперішній час горили і шимпанзе (родина людиноподібних мавп). Виділяють в окрему область Мадагаскар, для якого характерні нижчі мавпи – лемури і найпримітивніші комахоїдні – тенреки, або щетинисті їжаки.
Фауна Східної (Індо-Малайської) області (Південно-Східна Азія і Малайський архіпелаг) різноманітна, хоча і має багато спільних елементів з сусідніми областями. Тільки тут зустрічаються шерстокрили, тупайї і довгоп’яти, з людиноподібних мавп – гібони і орангутанги. З гризунів різноманітні білки, летяги і мишачі. Багато кажанів. Характерний індійський слон. Різноманітні парнокопитні (свині, бики, олені). Непарнокопитні представлені тапірами і носорогами. З хижих найбільш своєрідні котячі (тигр і дрібні коти) і панди.
В досить великій за площею Голарктичній області, куди входять Північна Африка, Європа, більша частина Азії і Північна Америка, ендемічний лише ряд зайцеподібних. З комахоїдних найбільш характерні кроти, їжаки, землерийки. Рукокрилих мало. Примати практично відсутні. З різноманітних гризунів характерні бабаки, ховрахи, бобри, тушканчики. Відносно мало парнокопитних (лосі, олені, козли, барани). З хижих найбільш широко поширені кунячі і собачі (вовк, лисиця, песець). Ссавці Євразії (Палеарктика) і Північної Америки (Неоарктика) часто представлені однаковими або близькими видами (ведмеді, рисі, вовки, бобри, зайці, зубр і бізон і т.п.). Тільки в Палеарктиці зустрічаються хохуля, кабан, верблюди, кінь Пржевальського і ряд інших видів, тільки в Неоарктиці – опосуми, єноти, скунси, гофери, вилорога антилопа і ін. Відносна бідність фауни Голоарктики визначається її північним положенням.
У морях середніх і високих широт обох півкуль поширені ластоногі. Китоподібні заселяють всі моря і океани; кити більш різноманітні в середніх і високих широтах, дельфіни – в тропічній зоні.
Положення ссавців в біоценозах. Заселяючи практично всю Землю, ссавці займають у біоценозах суші і морів екологічні ніші як первинних споживачів рослинних кормів (гризуни, зайцеподібні, копитні, хоботні, сирени, лінивці, дамани, значна частина приматів), так і м’ясооїдних, яких звичайно поділяють на „мирних” (комахоїдних, планктоноїдних), що поїдають дрібних тварин, і „хижаків”, нападаючих на велику за розмірами активну здобич. До перших відносяться комахоїдні, рукокрилі (за винятком нектарососів і плодоїдних криланів), неповнозубі, трубкозубі і вусаті кити; до других – зубаті кити, ластоногі і хижі. Змішане харчування зустрічається серед гризунів, хижих і приматів. Займаючи верхні щаблі ланцюгів живлення, ссавці, як і птахи і комахи, мають вплив на еволюцію рослин. Прикладом виступають забарвлення, форма, смак плодів і запах квітів, що привертають увагу тварин з високорозвиненими органами чуття, розвиток колючок і поява алкалоїдів, що зменшують поїдання рослин і т.д. Існують пристосування для прикріплення насіння і спор до тварин – поширювачів.
З діяльністю ссавців пов’язана стійкість біогеоценозів. Вимирання або винищення риючих гризунів (полівок, піщанок) в пустелях і степах супроводжується розростанням деяких видів рослин (багатолітніх, дернових злаків) і знищенням або скороченням кількості видів дводольних або однодольних ефемерів, що збіднює, а інколи і згубно діє на пасовиська. Помірний випас копитних сприяє підтриманню високої продуктивності травостою пасовиськ, а виключення випасу, як і перевипас, ведуть до знищення степового рослинного покриву. Все це свідчить про тісні взаємозв’язки рослин і тварин в біогеоценозах. За кількістю і біомасою серед хребетних у водоймах ссавці поступаються рибам, а на суші звичайно займають перше місце. В роки зростання чисельності дрібні ссавці, особливо полівки і миші, можуть досягати щільності 100-200 звірків на 1 га, а у скупченнях їх число піднімається до 1-5 на 1 м2. В роки депресії чисельності вони зберігаються тільки в стаціях переживання, всього на 1-10% придатної площі, заселяючи її зі щільністю менше 10-15 звірків на 1 га. Великі за розмірами копитні і ластоногі під час міграцій або в місцях розмноження також утворюють великі скупчення на обмеженій території. Решта часу вони тримаються невеликими групами по 1-5 особин на 1000 га. Щільність населення великих за розмірами хижаків (котячі, собачі) у сприятливих умовах не перевищує 1-3 на 1000 га. Дрібні хижі (кунячі) мають у 2-3 рази вищу щільність.
Значення ссавців для людини
Корисні ссавці багаточисельні і різноманітні. Серед давно приручених домашніх тварин вони займають більше 60% (15 видів). До них потрібно додати хутрових звірів, що розводять у клітках, і які знаходяться на ранній стадії доместикації (приблизно 20 видів), а також багаточисельних лабораторних тварин (щурі, миші, морські свинки і багато інших). Розведення у неволі супроводжується виведенням нових порід цих тварин. Кількість порід собак сягає 200 і продовжує збільшуватися; у кролів відомо більше сотні порід; серед лабораторних тварин збільшується число генетично чистих ліній мишей і щурів. Продовжується одомашнення нових видів: лося як транспортної тварини, оленя-марала для отримання пантів; нових хутрових звірів і лабораторних тварин. Для покращення існуючих і отримання нових порід використовується гібридизація домашніх тварин з дикими видами.
Мисливсько-промислові тварини дають чималий прибуток в багатьох районах. Успішна охорона їх запасів і відновлення чисельності майже знищених хижацькою експлуатацією соболя, бобра, калана, котиків і інших тварин збільшує мисливські багатства України і суміжних країн; успішна акліматизація американських гризунів – ондатри і нутрії – також збільшило їх. Добування соболя, куниць, білки, песця, лисиці і інших хутрових звірів разом з розведенням найбільш цінних хутрових звірів в неволі забезпечують потреби в хутровині. Основними промисловими видами лісової зони залишаються білка, соболь, куниця, горностай, лисиці і зайці. У зв’язку з ростом чисельності цінних кунячих білка поступилась їм першим місцем (по собівартості здобутих шкірок). В тундровій зоні основне значення зберігають песець і заєць-біляк, в степах і пустелях – лисиці, зайці, дрібні кунячі, ховрахи; в долинах річок – ондатра, водяна полівка, видра, а на півдні нутрія; у гірських районах важливе значення мають бабаки. Інші види (ведмеді, котячі і ін.) складають лише незначну долю здобутої хутровини. Дикі копитні – важливе джерело розв’язання потреб в м’ясі і шкірі: північні олені в тундрі, лосі і олені в лісовій зоні, сайгак в степах і напівпустелях. Особливе місце займає морський звіробійний промисел або на ліжбищах (котики), куди звірі приходять розмножуватись, або з морських суден (китобійний і тюленячий промисли). У першому випадку відлов з перевагою молодих самців. Морський китобійний промисел регулюється міжнародними угодами, поки що не завжди гарантуючими збереження основного поголів’я тварин, чисельність яких помітно зменшується. Продуктами морського промислу служать не лише шкіра (хутро) тварин, а і їх жир і деякі інші цінні продукти (наприклад, спермацет кашалотів).
У перелік шкідливих тварин включають хижаків, що нападають на домашніх тварин і людину, шкідників лісу і сільськогосподарських рослин, носіїв і поширювачів захворювань людини і домашніх тварин. Напади великих хижих (тигр, лев, леопард, вовки, гієни, ведмеді і ін.) на людину досить рідкісні і звичайно це робиться особинами, що втратили здатність добувати природну здобич. Таких „людоїдів”, безумовно, потрібно знищувати. Напад вовків на людину майже завжди пов’язаний з захворюванням вовків сказом. Необхідно обмежувати чисельність вовків і інших хижаків у місцях, де ті нападають на сільськогосподарських тварин, однак не слід перебільшувати їх шкідливість. Нерідко до цього вовків примушує зменшення чисельності копитних – їх природного корму. Різке скорочення чисельності великих хижих змушує приймати заходи щодо їх охорони: заборонено відстрілювати білого ведмедя, тигра, снігового барса.
Посівам, насадженням і пасовиськам шкодять гризуни і зайцеподібні. Їх шкідливість збільшується і тим, що багато мишей і полівок здатні до масових розмножень. Садам шкоду наносять соні і миші. Винищення шкідників сільського і лісового господарства ведеться переважно отрутохімікатами, що дозволяє зменшити їх чисельність, але нерідко отруює навколишнє середовище і не усуває небезпеки повторних масових розмножень.
Багато ссавців є носіями і поширювачами небезпечних захворювань людини і домашніх тварин, а також на ссавцях годуються переносники цих захворювань: кліщі, блохи, воші, москіти. Бабаки, ховрахи, піщанки, деякі полівки, миші і інші гризуни (всього близько 200 видів) є носіями і поширювачами небезпечного мікроба, що спричиняє чуму. Водяні щурі (всього більш 60 видів) поширюють туляремію. Вірусні інфекції, особливо енцефаліти, що передаються кліщами, пов’язані з дрібними ссавцями (личинково-німфальні фази кліщів) і великими ссавцями – хижими, копитними (дорослі кліщі). Близьке коло носіїв існує у геморрагічної лихоманки, кліщового сипного і зворотного тифів. Пустельні гризун є господарями шкірного лейшманіозу, а собачі – вісцерального лейшманіозу. Без участі переносників ссавці зберігають і передають інфекцію сказу (хижі, деякі кажани), еризипелоїда, лептоспірозів і листеріоза (полівки, щурі, миші, комахоїдні, хижі і копитні). Багато з цих інфекцій мають природні вогнища, тобто постійно існують у природі. Людина може захворіти контактуючи з хворою твариною або зараженою переносником (блохою, кліщем і т. п.), потрапивши на територію вогнища.