

Статеві органи і розмноження
Ссавці вирізняються високим ступенем турботи про потомство на всіх етапах його розвитку. Їм властивий розвиток в утробі самки, протягом якого зародок росте за рахунок речовин тіла матері, які він отримує через особливий орган – плаценту. Лише однопрохідні ссавці відкладають великі, багаті жовтком яйця.
У всіх ссавців після народження молодої тварини зв’язок з матір’ю не припиняється, так як дитина харчується молоком. Однак, і після завершення вигодовування молоком зв’язки між батьками і потомством, як правило, деякий час зберігаються. Це забезпечує можливість навчання молодняку – передачі накопиченого батьками індивідуального досвіду потомству. Такий зв’язок поколінь дозволяє накопичувати і сумувати популяційно-видовий досвід, має назву „сигнальної спадковості” або „сигнальної передачі інформації”. Це явище широко поширене серед птахів і особливо характерне для ссавців. Якісно іншого рівня набуває це явище у людини з її розумом і другою сигнальною системою, основою діяльності якої є виробничо-суспільні відносини.
Статеві органи ссавців більш складно збудовані , ніж у інших амніот (рис. 21). Сім’яники парні, розташовані у задній частині черевної порожнини (клоачні, деякі комахоїдні, неповнозубі, хоботні, китоподібні, сирени, носороги) або перемістились в мошонку – шкірястий виріст, що сполучається з порожниною тіла пахвинним каналом (самчасті, хижаки, копитні, примати). До сім’яника прилягає витягнуте по осі тіло – придаток сім’яника (epididymis) – скупчення вивідних проток сім’яника (залишок переднього відділу мезонефричної нирки). Від придатка відходить вольфів канал – сім’япровід, що впадає коло основи статевого члена в його сечостатевий канал. Перед впаданням в цей канал сім’япроводи утворюють дольчасті розширення – сім’яні міхурці (vesicula seminalis), секрет яких бере участь в утворенні сперми. У гризунів їх секрет виділяється після викидання сім’я в статеві шляхи самки, і, застигаючи, закупорює їх після копуляції.
Б
Рис. 21 Чоловічі статеві органи ссавців. А – сім’яники, розташовані в черевній порожнині; Б – сім’яники в мошонці:
1 – нирка, 2 – залоза надниркова; 3 – сім’яник; 4 – придаток сім’яника; 5 – сім’япровід; 6 – сім’яний пухирець, 7 – пряма кишка, 8 – сечовий міхур, 9 – передміхурова залоза, 10 – куперові залози; 11 – пеніс; 12 – препуціальні залози; 13 – пахвинний канал; 14 – стінка живота; 15 – стінка мошонки.
іля основи статевого члена лежить передміхурова залоза (prostata), секрет якої утворює рідке середовище сперми. Статевий член (penis) має печеристі тіла, що оточують сечостатевий канал; наповнення їх порожнин кров’ю забезпечує пружність копулятивного органу. У деяких ссавців (хижі, ластоногі, гризуни та ін.) його міцність збільшується спеціальною кісткою (os penis). З статевим членом пов’язані також куперові і препуціальні залози. Перші беруть участь в утворенні рідкої частини сперми (ейякулята), а другі виділяють пахучий секрет, що сприяє зустрічі особин протилежної статі і створенню статевого збудження партнерів.Парні яєчники самок завжди лежать в порожнині тіла і прикріплені до спинної стінки черевної порожнини брижами. Розмір яєчників помітно менший, ніж у решти хребетних. Парні яйцепроводи, гомологічні мюллеровим каналам, відкриваються в порожнину тіла біля яєчників лійками, що вистелені миготливим епітелієм. Верхній тонкий і звивистий відділ яйцепроводу називається фаллопієвою трубою, а нижній – розширений, з м’язовими стінками – маткою. У клоачних матки відкриваються в сечостатевий синус клоаки; у решти ссавців клоака зникає. У сумчастих нижня частина яйцепроводів перетворюється в парні піхви, які інколи зростаються кінцями і відкриваються в переддвір’я піхви (сечостатевий синус). У плацентарних ссавців нижні ділянки яйцепроводів зливаються в непарну піхву (vagina). Якщо кожна матка відкривається у піхву самостійним отвором, її називають подвійною (багато гризунів, слони та ін.); при двороздільній матці права і ліва матки зростаються кінцями і відкриваються у піхву загальним отвором (гризуни, деякі хижаки, свині), а при дворогій – зростаються приблизно наполовину своєї довжини (комахоїдні, хижі, копитні, китоподібні). У частини рукокрилих і приматів права і ліва матки зливаються в єдину просту матку, в яку відкриваються права і ліва фаллопієві труби (рис 22).
П іхва
переходить в короткий розширений
сечостатевий синус (переддвір’я піхви),
в який відкривається і протока сечового
міхура. У стінці переддвір’я піхви
розташований невеликий виріст з
печеристими тілами – клітор, гомологічний
копулятивному органу самців. Переддвір’я
піхви відкривається назовні щілиноподібним
сечовим отвором, обмеженим згортками
шкіри – статевими губами. Розташовані
тут шкірні залози виділяють пахучий
секрет.
іхва
переходить в короткий розширений
сечостатевий синус (переддвір’я піхви),
в який відкривається і протока сечового
міхура. У стінці переддвір’я піхви
розташований невеликий виріст з
печеристими тілами – клітор, гомологічний
копулятивному органу самців. Переддвір’я
піхви відкривається назовні щілиноподібним
сечовим отвором, обмеженим згортками
шкіри – статевими губами. Розташовані
тут шкірні залози виділяють пахучий
секрет.
Я
Рис. 22. Схема еволюції яйцепроводів у ссавців. І – клоачного; ІІ – нижчого сумчастого (опосум); ІІІ – вищого сумчастого (кенгуру); ІV – плацентарного ссавця з подвійною маткою; V – те ж, з дворогою маткою; VI – те ж, з простою маткою:
1 – фаллопієва труба, 2 – матка, 3 – піхва, 4 – сечостатевий синус, 5 – сечовий міхур, 6 – клоака, 7 – пряма кишка
йцеві клітини – оогонії – утворюються в яєчниках із зачаткового епітелію; вони ростуть і перетворюються в ооцити; кожний ооцит оточується фолікулярними клітинами, утворюючи первинний фолікул. До часу розмноження ооцит починає рости, накопичуючи жовток, а його оболонка розростається і між нею і яйцеклітиною – ооцитом утворюється заповнений рідиною простір. Фолікул перетворюється у граафів пухирець, що видається на поверхні яєчника. Потім оболонка пухирця лопається і яйцеклітина випадає в порожнину тіла (відбувається овуляція), звідки переходить у фаллопієву трубу, де і запліднюється. Утворений на поверхні яєчника рубець заповнюється кров’яним згустком, проростає лютеїновими клітинами, що містять жовтий пігмент і володіють гормональними функціями. Так у яєчнику утворюється жовте тіло, що виділяє гормон прогестерон. При заплідненні яйцеклітини і настанні вагітності розвивається жовте тіло, а прогестерон стимулює імплантацію зародка в стінку матки, одночасно затримуючи утворення нових граафових пухирців і овуляцію до закінчення вагітності.Оболонки зародка сумчастих ссавців в період розвитку лише прилягають до стінки матки, викликаючи частковий гістоліз її поверхневих шарів. У частини видів утворюється зачаток зародкової частини плаценти шляхом зростання зовнішньої стінки алантоїса з серозою, але без утворення ворсинок, що проникають в товщу стінок матки. Харчування зародка відбувається за рахунок жовтка яйця і секреторних виділень клітин стінки матки. У плацентарних ссавців виникає справжня плацента: зрослі ділянки серози і зовнішньої стінки алантоїса утворюють хоріон (зародкову частину плаценти), вирости (ворсинки) якого входять в стінку матки; в цьому місці остання розрихлюється і утворює материнську частину плаценти. При цьому посилюється гістоліз тканин матки, і кровоносні судини зародка тісніше стикаються з судинами материнського організму. Це полегшує харчування зародка за рахунок речовин крові матері (гемотрофне харчування). Утворення плаценти називається імплантацією зародка. Плацента може мати різну будову: її називають дифузною, коли ворсинки розміщені рівномірно по всьому хоріону, або часточкова, коли вони зібрані в окремі групи, або дискоїдальною, коли ворсинки зосереджені в одному місці, утворюючи диск і т. д. Ступінь зв’язаності зародкової і материнської плацент неоднаковий і за цією ознакою розрізняють відпадаючу і невідпадаючу плаценти. У першому випадку зв’язок настільки глибокий, що при родах частина стінки матки відривається і виводиться назовні у вигляді посліду.
У одних видів дитинчата ссавців народжуються безпорадними, в інших – здатними до активних дій. Новонароджені сумчасті особливо дрібні. Вони недорозвинені і доношуються у спеціальних сумках. Тому у сумчастих тривалість вагітності невелика: опосум – 12 днів, гігантський кенгуру – 38―40 днів. Чим більші розміри тварини, тим, зазвичай, триваліший період вагітності. Залежить він і від екології виду. Комахоїдні народжують недорозвинених малят після 13―19 днів вагітності; у той же час у кажанів народжуються більш самостійні малята, вагітність триває 54―73 дні; у тварин, що народжують малят в норах з теплими гніздами, вагітність нетривала (хатня миша – 18 днів, сіра полівка – 16―23; ондатра – 25―26 дні); більша вона у великих за розмірами норників-бабаків (30―40 днів). У звірів, які не мають добре влаштованих сховищ, вагітність триваліша: у нутрії близько 130 днів, леопарда 120, бурого ведмедя 200 днів. Найтриваліша вагітність у тварин, малята яких відразу після народження слідують за самкою: у копитних 150 (свині) – 290 (бики) днів; у білого носорога 540 днів, у слонів більше 600 днів; велика вона у ластоногих (морж 200 днів) і китоподібних (270―365 днів).
Інколи вагітність продовжується за рахунок латентного періоду, коли зародок на деякий час призупиняє свій розвиток. Така діапауза дозволяє відкласти роди до сприятливого періоду. Вона встановлена для, так званих, діестральних видів, тобто тих, що мають два період статевої активності. Так, у борсука перша тічка проходить в липні-серпні, а друга – в жовтні. Запліднені в липні яйця знаходяться в спокої до глибокої осені, а запліднені в жовтні розвиваються без діапаузи; дитинчата народжуються весною. У горностая запліднені під час весняної тічки самки приносять виводки через два місяці, а запліднені в серпні-вересні – тільки через 8―9 місяців. У козулі запліднені в першу тічку (липень-серпень) яйця сповільнено розвиваються до грудня, а запліднені під час осінньої тічки (листопад-грудень) - затримки розвитку не мають. У куниці і соболя справжня тічка припадає на середину – кінець літа, а друга (несправжня) – на початок весни. Вона пов’язана з імплантацією діапаузованих яєць (роди відбуваються навесні).
Швидкість постнатального (після народження) розвитку ссавців на перших стадіях пов’язана зі складом молока, яким мати вигодовує малят. Молоко ссавців має всі необхідні для розвитку речовини: білки, жири, вуглеводи, вітаміни, солі. Швидкість розвитку тим вища, чим більший в молоці вміст білків і жирів.
Розмноження ссавців ритмічне, т.т. впорядковане в часі. Його можливості визначаються тривалістю статевих циклів, які розпочинаються після статевого дозрівання. Терміни статевого дозрівання широко варіюють. Найбільш плодовиті дрібні тварини: полівки досягають статевої зрілості у віці 1, 5―2 місяці, миші – 2―3 місяці, ондатра – 5 місяців, зайці – близько року. Більші за розміром звірі починають розмножуватися пізніше: вовки, куниці, лисиці і соболі – на 2 році життя, тигри, ведмеді, багато видів тюленів і китоподібних – на 3―4 році, олені – на 2-4 році життя, носороги і слони – в 10―15 років. Статева зрілість у великих за розміром мавп настає у віці 10―12 років. Частота розмноження і його періодичність пов’язані з величиною смертності і тривалістю життя. Висока плодючість – важливе пристосування дрібних ссавців, які фізіологічно нестійкі і мають багаточисельних і добре озброєних ворогів. Вона компенсує високу їх загибель. Багато гризунів потенційно здатні розмножуватися протягом цілого року, навіть в суворих умовах Арктики. Так, зимове розмноження існує у норвезького лемінга. Багато разів його реєстрували також у сибірського лемінга. Зимове розмноження спостерігали у мишей і полівок, піщанок і хом’яків в лісовій, степовій і пустельних зонах. Щурі і хатні миші в людському помешканні також здатні розмножуватися протягом усього року. Велика амплітуда коливань плодючості і смертності пояснює дуже мінливу чисельність перерахованих вище тварин. Решта видів, як правило, розмножуються в певний період року. Тривалість репродуктивного циклу залежить від тривалості вагітності. Роди завжди припадають на сприятливий для вигодовування молоді період, як правило, на весну. В період розмноження овуляція і тічка самок циклічно повторюються до запліднення. Такі статеві естральні цикли звичайно складаються з 4 фаз: передтічка (prooestrus), тічки (oestrus), післятічкового періоду (metaoestrus) і періоду спокою (disoestrus). Естральний цикл у дрібних гризунів триває близько 6 днів, у собак – 9―14, у копитних (кіз, овець, корів) – 17―24 дні.
Готовність до розмноження і приуроченість часу парування до певного періоду забезпечуються складним регуляторним механізмом, який діє за сигналами зовнішнього середовища. В помірних і високих широтах таким сигналом є зміна довжини світлового дня. Для видів з весняною тічкою сигнальне значення має збільшення довжини дня (багато тварин), для копитних, які утворюють пари восени – його скороченні, а для вовків – короткий зимовий день. Зміна світлового режиму впливає на нейрогіпофіз, який стимулює виділення передньою долею гіпофіза гонадотропних гормонів, що в свою чергу, впливають на гонади. В низьких широтах значення сигнального фактора можуть мати й інші періодичні явища природи, що передують наближенню сприятливого для розмноження періоду (в пустелях і саванах – опади, в тропічних лісах – дозрівання основних кормових рослин, або їх плодів і т. д.). Навіть у тропічних лісах з їх постійним теплим і вологим кліматом, розмноження ссавців зазвичай сезонне, хоча у різних видів припадає на певні періоди року.
Загальна плодючість ссавців, завдяки високому рівню і різноманітним формам турботи про потомство, невелика. У дрібних гризунів з їх високою смертністю виводок може сягати 12 малят (звичайно 5―6), а розмноження триває протягом усього року (до 6 приплодів на рік). Зайці і білки приносять 2―3 приплоди в рік, кожен з яких зазвичай складає від 3 до 8 малят (до 12). Вовки, лисиці, коти, соболі, куниці, горностаї розмножуються раз на рік і мають виводки з 3―6 малят. Раз на рік народжують 1―2 малят порожнисторогі, олені, тюлені і дельфіни. Слони, вусаті кити, тигри, деякі великі котячі розмножуються раз на 2―3 роки, народжуючи зазвичай 1―2 малят. Плодючість змінюється не лише в залежності від зовнішніх умов (забезпеченість їжею, погода), але і від щільності (чисельності) популяції: при її зростанні збільшується доля ялових (не беруть участь у розмноженні) самок і зменшується величина виводків.
Тривалість життя слонів дорівнює 70―80 років, собачих – 10―15, великих котів і китів – 30―40, білячих – 8―10, дрібних гризунів – 1―3 роки.
Хижаки і гризуни в період розмноження утворюють пари або складні сім’ї (парцели гризунів, прайди левів). Такі групи з декількох самців, самок і молодих особин краще забезпечують виховання останніх. Вони часто зберігаються і після періоду розмноження. У стадних копитних, ластоногих, китоподібних в період парування утворюються тимчасові „гареми”, які складаються з декількох самок на чолі з самцем-домінантом. Після парування самки з молодняком інколи утворюють окремі угруповання. Окремо тримаються поза періодом розмноження самці і самки кажанів.
Ендокринна система ссавців принципово подібна до гормональної системи решти хребетних тварин. Відмінності полягають не стільки в складі і числі гормонів, скільки в посиленні їх активності і спеціалізації. Особливо це стосується гормонів, що беруть участь у здійсненні статевого циклу, вагітності і лактації (у самок – естрогени, пролактини, у самців – андрогени). Тісний взаємозв’язок гормональної діяльності ендокринних залоз між собою і центральною нервовою системою забезпечує узгоджене протікання всіх життєвих процесів організму (в тому числі інтенсивності і характера метаболізму, їх „підлаштування” до змінних умов зовнішнього середовища). Як і в птахів, розвиток вторинних статевих ознак у ссавців визначається не тільки статевими відмінностями в характері генотипу, але і гормонами статевих залоз.
Н ервова
система і нервова діяльність
ервова
система і нервова діяльність
Головний мозок ссавців, зберігаючи спільні з іншими хребетними риси, відрізняється принциповими особливостями, що виділяють його в особливий кортикальний тип.
Рис. 23. Пошарове розташування нервових клітин в корі півкуль переднього мозку ссавців.
У головному мозку ссавців найбільшої величини і складності досягає передній мозок. Більша частина мозкової речовини зосереджена в корі півкуль, тоді як смугасті тіла відносно невеликі. Кора переднього мозку формується шляхом розростання нервової речовини стінок бічних шлуночків. Утворене в такий спосіб мозкове склепіння має назву вторинне склепіння або неопалліум (neopallium); його зачатки з’являються у амфібій, більш помітні у рептилій і птахів. Неопалліум складається з нервових клітин і безм’якотних волокон (сіра речовина мозку). Обидві півкулі з’єднані між собою комісурою з білих (мієлінових) волокон. Комісура має назву мозолисте тіло. Тіла нейронів в корі півкуль розташовані пошарово, утворюючи своєрідні „екранні структури” (рис. 23). Така організація мозку дозволяє просторово відображати зовнішній світ на основі інформації, що надходить від органів чуття. Екранні структури характерні для найважливіших мозкових центрів ссавців, тоді як в інших хребетних вони зустрічаються рідко, переважно в зорових центрах. Нова кора великих півкуль служить центром вищої нервової діяльності, який координує роботу інших відділів мозку (рис. 24). Лобові долі відповідають за спілкування тварин, в тому числі акустичне; у людини вони пов’язані з мовою, тобто з другою сигнальною системою. Кора
майже всіх ссавців утворює більшу чи
меншу кількість борозен, які збільшують
її поверхню. У найпростіших випадках є
тільки одна сільвієва борозна, яка
відділяє лобову долю від вискової;
пізніше з’являється роландова борозна,
яка розділяє лобову і потиличну долі
та ін. У приматів і зубатих китів кількість
борозен особливо велика. Неопалліум
ссавців в більшій мірі, ніж комплекс
кори середнього мозку птахів, забезпечує
вищу нервову діяльність, нагромаджуючи
сліди поодиноких збуджень та їх поєднань,
збагачуючи так з вану
оперативну пам’ять. Це дає можливість
на її основі вибирати в новій ситуації
оптимальне рішення. Найчастіше вони
представляють нові комбінації вже
відомих елементів. Виникнення у ссавців
вищих асоціативних центрів – нової
кори – не вело до ліквідації центрів,
що керують інстинктивними актами, а
лише підпорядкувало останні вищому
контролю.
вану
оперативну пам’ять. Це дає можливість
на її основі вибирати в новій ситуації
оптимальне рішення. Найчастіше вони
представляють нові комбінації вже
відомих елементів. Виникнення у ссавців
вищих асоціативних центрів – нової
кори – не вело до ліквідації центрів,
що керують інстинктивними актами, а
лише підпорядкувало останні вищому
контролю.
Рис. 24. Функціональні центри кори головного мозку землерийки (А) і людини (Б):
1 – руховий центр; 2 – центр шкірно-м'язової чутливості; 3 – зоровий центр; 4 – слуховий центр; 5 – нюхова цибулина; 6 – нюхові долі; 7 – склепіння середнього мозку; 8 – мозочок; 9 – лобова доля.
Інші частини переднього мозку мають відносно менші розміри, але також зберігають своє значення. Нюхові долі розташовані в передній частині знизу (нюхові цибулини і передні базальні ядра). Смугасті тіла виконують функцію регуляції інстинктивних реакцій під контролем кори півкуль.
Рис. 25 Схема зв’язків між відділами головного мозку ссавців:
1 – нюховий нерв; 2 – нюхова цибулина; 3,4 – базальні ганглії; 5 – неопалліум; 6 – дорзальна частина зорового горба; 7 – його вентральна частина, медіальне(8) і латеральне(9) колінчасте тіло; 10 – зоровий нерв; 11 – передні (зорові) і 12 – задні (слухові) горбики даху середнього мозку; 13 – ретикулярна формація середнього мозку; 14 – руховий пірамідальний тракт; 15 – слуховий нерв; 16 – слуховий центр довгастого мозку; 17 – чутливі нерви шкіри і м’язів голови; 18 – чутливі ядра стовбура мозку; 19 – чутливі нерви шкіри і м’язів тіла; 20 – чутливі (дорзальні) роги спинного мозку; 21 – чутливі ядра довгастого мозку; 22 – ретикулярна формація довгастого і спинного мозку; 23 – рухові ядра стовбура мозку; 24 – рухові нерви до м’язів голови; 25 – рухові (вентральні) роги спинного мозку; 26 – рухові нерви (до м’язів тіла).
 Середній
мозок малий; його дах поділений поперечними
борознами і носить назву чотиригорбикове
тіло. Його передні горби утворюють слабо
виражену зорову кору, задні підконтрольні
передньому мозку і служать слуховими
центрами (Рис. 25). Мозочок великий і
складається з серединного черв’ячка
і прилеглих до нього півкуль з бічними
придатками. Як і в інших ссавців, з
мозочком пов’язані підтримка м’язового
тонусу, пози, рівноваги, рухів частин
тіла. Тісні зв’язки мозочка з корою
півкуль свідчать про існування над ним
вищого контролю.
Середній
мозок малий; його дах поділений поперечними
борознами і носить назву чотиригорбикове
тіло. Його передні горби утворюють слабо
виражену зорову кору, задні підконтрольні
передньому мозку і служать слуховими
центрами (Рис. 25). Мозочок великий і
складається з серединного черв’ячка
і прилеглих до нього півкуль з бічними
придатками. Як і в інших ссавців, з
мозочком пов’язані підтримка м’язового
тонусу, пози, рівноваги, рухів частин
тіла. Тісні зв’язки мозочка з корою
півкуль свідчать про існування над ним
вищого контролю.
Довгастий мозок дає початок більшості головних нервів (V―XII). У ньому розташовані центри дихання, роботи серця, травлення і т.п. З боків порожнини четвертого шлуночка відокремлюються пучки нервових волокон, які йдуть до мозочка і утворюють його „задні ніжки”. Нервові тракти зв’язують довгастий мозок зі спинним.
Як і в інших хребетних, відносні розміри головного мозку збільшуються при зменшенні розмірів тіла і збільшенні напруження терморегуляції. Так, у великих комахоїдних маса головного мозку складає близько 0,6% маси тіла, а у дрібних – до 1,2, у великих китоподібних – близько 0,3%, а в дрібних – до 1,7%. Маса мозку приматів складає 0,6―1,9% від маси тіла, в людини – близько 3%. У всіх ссавців маса переднього мозку перевищує масу інших відділів головного мозку: в різних групах вона складає 52―72% загальної маси головного мозку; у приматів цей показник зростає до 76―80%, а в людини – до 86%.
Співвідношення мас головного і спинного мозку максимальне у людини (45:1), високе у приматів і китоподібних (10―15:1), нижче у хижаків, комахоїдних (3―5:1) і копитних (2,5:1). У рептилій воно завжди менше 1. Спинний мозок з участю провідних шляхів (біла речовина) пов’язаний з рушійним центром кори півкуль, який здійснює вищий контроль над руховими актами і управління складними рухами.
Спинні стовбури білої речовини складаються з висхідних до головного мозку волокон, що несуть імпульси від органів чуття і ентерорецепторів (аферентна інформація). У черепних стовбурах переважають волокна, які несуть імпульси від мозку до м’язів та інших виконавчих органів (еферентна інформація). Контроль вищих центрів головного мозку над роботою спинного мозку досягає у ссавців найбільшого рівня.

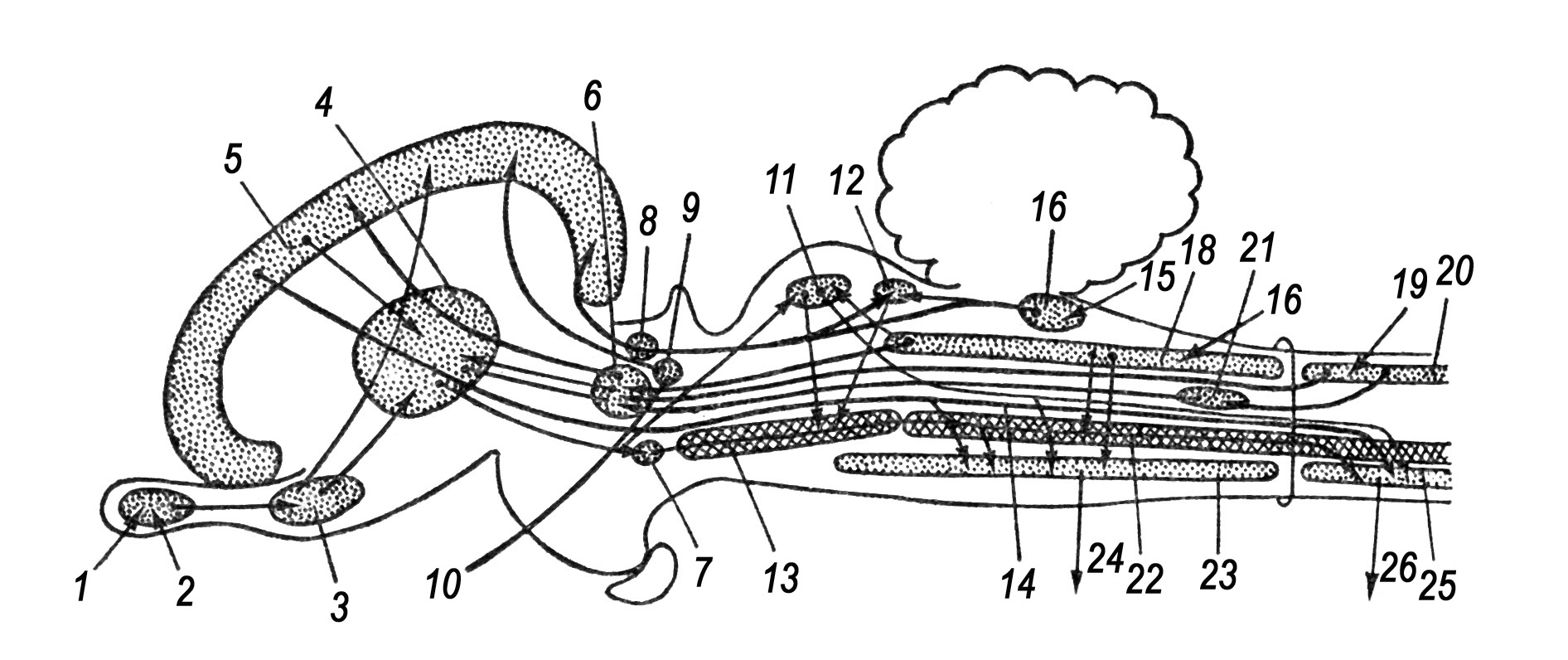
Рис. 26. Схема вегетативної нервової системи ссавців:
1 – парасимпатичні центри в середньому і довгастому мозку; 2 – те ж в крижовому відділі спинного мозку; 3 – симпатичний стовбур; 4 – шийні вузли; 5 – зірчастий вузол; 6 – черевний вузол; 7 – передній і 8 – задній брижові вузли; 9 – тазовий вузол; 10 – око; 11 – слинні залози; 12 – щитовидна залоза; 13 – легеня; 14 – серце; 15 – печінка; 16 – шлунок; 17 – селезінка; 18 – підшлункова залоза; 19 – нирка; 20 – надниркова залоза; 21 – тонка і 22 – товста кишка; 23 – сечовий міхур; 24 – матка; 25 – пряма кишка. Темні лінії – іннервація парасимпатичної системи, пунктирні – іннервація симпатичної системи.
Ссавці мають 12 пар головних нервів; розвивається XІ пара – додаткові нерви (n. acсessorius). Крім іннервації основних органів чуття (нюх, зір, слух) і м’язової системи, головні нерви беруть участь в утворенні вегетативної нервової системи, яка контролює, так звані, вегетативні процеси, що не підпорядковується вольовому контролю. Парасимпатична нервова система утворена черепними нервами довгастого мозку і спинномозковими нервами крижового відділу. Симпатична складається з нервових вузлів спинномозкових нервів шийного, грудного і поперекового відділів хребта (рис. 26). Основні системи органів мають закінчення обох систем.
Паралельна іннервація пояснюється протилежно направленим впливом. Якщо імпульси однієї з них мають збуджуючий вплив на функцію органа, то імпульси іншого органа звичайно гальмують їх. Антагоністичний вплив, удосконалюючи регуляцію, значно розширює здатність переносити пригнічені або дуже збудливі зовнішні впливи (стрес), збільшуючи шанси виживання організму у широкому діапазоні умов.