

Ряд Безногі – Apoda
Д аний
ряд включає єдину родину червуги –
Caеcilidae. Нараховує , близько 60 видів,
ззовні нагадують великих червів або
змій з довжиною тіла 30―120 см.
Характеризуються видовженим червоподібним
тілом з кільцевими перетяжками, без
парних кінцівок із хвостом (рис. 102). Усі
безногі, за винятком представників
двох південноамериканських родів,
ведуть підземний спосіб життя, риють
нори у зволоженому ґрунті і в рослинній
підстилці. Це є спеціалізована група
тварин: має видовжену форму тіла без
кінцівок та їх поясів; рот розташований
знизу; очі зачаткові, відсутня барабанна
перетинка, слуховий нерв рудиментарний;
добре розвинені органи нюху; запліднення
внутрішнє, клоака вивертається, виконуючи
роль копулятивного органу.
аний
ряд включає єдину родину червуги –
Caеcilidae. Нараховує , близько 60 видів,
ззовні нагадують великих червів або
змій з довжиною тіла 30―120 см.
Характеризуються видовженим червоподібним
тілом з кільцевими перетяжками, без
парних кінцівок із хвостом (рис. 102). Усі
безногі, за винятком представників
двох південноамериканських родів,
ведуть підземний спосіб життя, риють
нори у зволоженому ґрунті і в рослинній
підстилці. Це є спеціалізована група
тварин: має видовжену форму тіла без
кінцівок та їх поясів; рот розташований
знизу; очі зачаткові, відсутня барабанна
перетинка, слуховий нерв рудиментарний;
добре розвинені органи нюху; запліднення
внутрішнє, клоака вивертається, виконуючи
роль копулятивного органу.
Живляться комахами, червами та іншими безхребетними ґрунту. Яйця розвиваються поза водою, лише остання стадія личинки пов’язана з водою. Заселяють Південну Америку, тропічну Африку та Південну Азію. Поширеним є рибозмій цейлонський – Ichtyophis glutinosus, який скручує тіло навколо своєї кладки, забезпечує її вологою. Деякі види живородні. Прогресивними особливостями представників безногих, як і хвостатих, є внутрішнє запліднення і турбота про потомство. Червуга кільчаста – Siphonops annulatus (рис. 102) поширена у Північній Бразилії, Перу, Еквадорі, досягає 40 см довжини, живе в ґрунті на берегах річок та озер. Самка відкладає 5 яєць у нори під пеньками.
Походження, еволюція земноводних. У верхньому девоні від прісноводних кистеперих риб відособились перші земноводні – іхтіостегіди – Ichthyostegalia. Можливо, вже у верхньому девоні вони дали початок двом підкласам земноводних – тонкохребцевим і дугохребцевих – пануючих у наземній фауні кам’яновугільного періоду та витісняючи прісноводних кистеперих риб із мілководних водойм. Палеозойських земноводних, незалежно від їх таксономічної приналежності, звичайно назвали стегоцефалами або панцирноголовими. Вони, були покриті суцільним панциром із покривних кісток (шкірних). В панцирі мались лише отвори ніздрів, очниць і тім’янного органа. У деяких стегоцефал тіло було покрите рибоподібною лускою, в інших шкірні окостеніння зберігались на нижній стороні тіла, утворюючи черевний панцир (захист черева при лазінні по суші). В багатьох видів тазовий пояс не з’єднувався з хребетним стовпом, а пояс передніх кінцівок деколи зберігав зв’язок з потиличним відділом черепа. Парні кінцівки були відносно слабкішими.
Дугохребцеві земноводні (підкл. Apsidospondyli) палеозойської ери об’єднуються в надряд лабіринтодонтів – Labyrinthodontia названих тому, що зовнішня поверхня їхніх зубів мала складну складчастість, на поперечному розпилі зуба утворюючи лабіринт емалевих петель. В цей надряд включають чотири ряди. В кінці девону ряд іхтіостегід дав початок багатьом групам земноводних; від нього ж відділилась гілка, що веде до рептилій. Ряд рахітомові – Rachitomi та відокремлений від нього в карбоні ряд стереоспондільних амфібій – Stereospondyli були представлені видами різного розміру (найбільші досягали 5 м довжини). Можливо, вони жили на берегах величезних рік і озер, здобич (переважно рибу) підчікували на мілководді. В частини видів, які мали велику голову тяжкий тулубовий хвіст був помітно вкорочений. Лабіринтодонти почали вимирати в кінці палеозою (у пермі) і лише небагато збереглося до середини тріасу. Ймовірно, від якихось примітивних рахітомових виникли стрибаючі безхвості земноводні – надряд Salientia. У відкладах нижнього тріасу знайдені залишки примітивних безхвостих (довжина біля 10 см тулубових хребців до 15 були короткі ребра, стегно довше за гомілку, можливо був короткий хвіст); їх об’єднує в ряд Proanura. Види, які зустрічалися десь зі середнього юра (тулубових хребців біля 10, гомілка не коротша стегна) відносять до ряду безхвості – Anurа, включаючи сюди і всіх сучасних безхвостих земноводних.
В кінці девону – початку карбону від якихось іхтіостегід або примітивних рахітомових відділився ряд антракозаври – Anthracosauria (або емболомери). За способом життя (досить крупні рибоїдні хижаки) і за загальними рисами будови вони були подібні на примітивних рахітомових, відрізнялись від них лише деякими деталями будови хребців, черепа (вирізки на його задньому краю, формування одного потиличного виросту. В середині карбону від них відділились сеймуріаморфми – Seymouriamorpha, еволюція яких проходила по шляху придбання все більшої наземності та рухливості. Ряд зоологів та палеонтологів, мають схильність відносити їх до класу земноводні виділяючи в підклас батрахозаврів – Batrachosauria.
В кінці девону від іхтіостегій відокремились тонкохребцеві земноводні – підклас Lepospondyli. Палеозойських лепоспондільних стегоцефал ділять на три ряди. Ряд мікрзаурія – Microsauria представлений дрібними формами (до 50 см довжини), зовні нагадували сучасних тритонів і саламандр. Від якихось, можливо, пермських мікрозаурій відділились нині існуючі ряди земноводних: хвостаті – Urodela (залишки відомі з середини крейдового періоду) і безногі – Apoda (викопні залишки поки що не виявлені). Ряд некриідіа – Nactridia об’єднує більш крупних (довжиною до 100 см) тритоноподібних земноводних, які вели переважно водний спосіб життя. Ряд аістопода – Aistopoda охоплює дрібних тварин (довжина 20-50 см) із змієподібним тілом і редукованими кінцівками. Вважалось, що вони вели водний спосіб життя; тепер висловлюється, можливо, більш обґрунтоване припущення, що утрата кінцівок була зв’язана з перебуванням в густих трав’яних заростях берегів водойм. Останні два ряди вимерли, не залишивши потомків.
Таким чином, адаптивна радіація палеозойських амфібій мала місце в кам’яновугільному і частково в перському періодах. В останньому періоді більшість груп вимерли і лише окремі форми деяких груп дожили до середини тріасу. Можливо таке швидке вимирання обумовлене біотичними, а не кліматичними причинами. В нижній пермі, як і в карбоні, на більшій частині суші переважав теплий і вологий клімат. Лише у верхній пермі і в тріасі, після вимирання більшості палезойських земноводних, він став більш сухим і жарким. Сеймуріаморфні і примітивні земноводні зберігали значні амфібійні риси, заселяли такі ж біотопи, що і земноводні. Маючи велику рухомість і, можливо, складну поведінку, сеймуріаморфи і відокремились від них у верхньому карбоні – пермі інші групи рептилій поступово витісняли (шляхом харчової конкуренції і прямого переслідування) земноводних із засвоєних ними місцях перебування. Довше уціліли земноводні з переважно водним способом життя, але і вони в тріасі відступили перед натиском рептилій, деякі форми котрих в цей час заселили водні біотопи, вторинно перейшовши до водного або напівводяного способу життя.
Анамнії (Anamnia) і Амніоти (Amniota)
Риби і земноводні відносяться до різних надкласів хребетних тварин, цим і підкреслюються принципові відмінності між ними. В той же час земноводні, як перехідний (амфібіотичний) клас, зберегли багато подібних з рибами рис. На основі цієї подібності надклас риби і клас земноводні об’єднують в групу анамній (не надаючи їй таксономічного значення); решта класів – плазуни, птахи, ссавці – формують групу наземних хребетних – амніот.
Для більшості анамній властиве зовнішнє запліднення, але для деяких хрящових риб і частини земноводних – внутрішнє. Яйце розвивається у воді, личинка, що вилупилася, веде водний спосіб життя і лише у небагатьох земноводних виникають пристосування, які забезпечують розвиток яйця поза водою.
Амніоти – первинноводні наземні хребетні тварини. Запліднення у них внутрішнє. У яйцекладних амніот яйце може розвиватися у повітряному середовищі. Амніоти, які вторинно перейшли до життя у водоймах всетаки відкладають яйця на суші (морські черепахи, крокодили та ін.), або у них розвивається живородіння (морські змії). Личинкова стадія відсутня, розвиток проходить без метаморфозу. Різниця між анамніями і амніотами проявляється у будові яєць, характері ембріонального розвитку та особливостях будови дорослих особин.
КЛАС Плазуни, або рептилії - Reptilia
Характеристика класу. Плазуни – перший клас справжніх первинноназемних хребетних (Amniota). Для них властиві крупні, багаті жовтком і білком яйця, покриті щільною пергаментною оболонкою. Запліднення лише внутрішнє. Ембріональний розвиток проходить в повітряному середовищі з утворенням зародкових оболонок – амніона, серози – і алантоїса, личинкова стадія відсутня. Молода тварина, яка вилупилась із яйця, відрізняється від дорослих лише розмірами. Суха шкіра плазунів майже позбавлена залоз. Зовнішні шари епідермісу роговіють: у шкірі утворюються рогові луски та щитки. Дихання лише легеневе. Утворюються повітроносні шляхи – трахеї і бронхи. Дихання здійснюється завдяки рухам грудної клітки. Серце трикамерне. Від розділеного неповною перегородкою шлуночка самостійно відходять три кровоносні стовбури: дві дуги аорти і легенева артерія. Сонні артерії, що забезпечують артеріальною кров’ю голову, відходять лише від правої дуги аорти. Велике і мале кола кровообігу повністю не розділені, але ступінь їх відокремлення вища, ніж в земноводних. Виділення і водний обмін забезпечують метанефричні (тазові) нирки, збільшуються відносні розміри головного мозку особливо за рахунок збільшення півкуль і мозочка.
Скелет повністю кістковий. Осьовий скелет (хребет) ділиться на п’ять відділів. Видовжена шия і два перші спеціалізовані шийні хребці (атлас і епістрофей) забезпечують високу рухливість голови. Череп має один потиличний виросток і добре розвинені покривні кістки; скроневі ямки і обрамлюючі їх кісткові дуги. Кінцівки наземного типу з інтеркарпальним і інтертарзальним з’єднаннями. Пояс передніх кінцівок з’єднується з осьовим скелетом через ребра, тазовий пояс зчленовується з поперечними відростками двох крижових хребців.
Плазуни заселяють різні наземні місця переважно в теплих, часто в помірних широтах; частина видів знову перейшла до водного способу життя. Загальний рівень життєдіяльності помітно вищий, ніж у земноводних. Хоча температура тіла непостійна і в значній мірі залежить від температури навколишнього середовища (пойкілотермія).
Особливості організації плазунів. Форма тіла плазунів більш різноманітна, ніж земноводних. Це пов’язано з способами пересування. Змієподібні плазуни (змії, безногі ящірки) втратили кінцівки, але успішно пересуваються в підстилці, в трав’яних заростях, в кронах дерев, а також добре плавають.
По твердому субстрату пересування особин досягається за допомогою кінцівок та підйому тіла над землею. Це дало можливість ящіркам (агами, круглоголовки) перейти до швидкого чотириногого пересування, а згодом до двоногого бігу. В тварин, які ведуть деревний спосіб життя видовжились задні кінцівки, які допомагають скакати по гілках, а розвиток шкіряної складки з боків тіла – планеруванні (літаючі дракони) (рис. 104).
В класі рептилій склались різні типи пересування по ґрунті, у воді та повітрі.
Пристосування до руху в різних середовищах було головною умовою прогресивного розвитку плазунів. Його основою слугувало використання всіх потенціальних можливостей механічної схеми, представленої центральним внутрішнім опорним скелетом, парними кінцівками і зв’язаною з ними мускулатурою.
П окриви.
Верхні шари багатошарового епідермісу
плазунів роговіють: клітини заповнюються
зернятками білка кератину, що витісняють
цитоплазму і ядро. Під цим мертвим шаром
розташований нижній мальпігієвий шар,
який складається із живих епідермальних
клітин, які розмножуються (рис. 103). За
рахунок розростання рогового шару
утворюються щитки, луски, рогові
зернятка, горбики, шипи, кігті. Під
роговими лусками у деяких видів плазунів
в мезодермальному шарі шкіри – коріумі
– залягають кісткові пластинки. У
черепах вони зливаються в кістковий
панцир, що приростає до хребта.
окриви.
Верхні шари багатошарового епідермісу
плазунів роговіють: клітини заповнюються
зернятками білка кератину, що витісняють
цитоплазму і ядро. Під цим мертвим шаром
розташований нижній мальпігієвий шар,
який складається із живих епідермальних
клітин, які розмножуються (рис. 103). За
рахунок розростання рогового шару
утворюються щитки, луски, рогові
зернятка, горбики, шипи, кігті. Під
роговими лусками у деяких видів плазунів
в мезодермальному шарі шкіри – коріумі
– залягають кісткові пластинки. У
черепах вони зливаються в кістковий
панцир, що приростає до хребта.
В мальпігієвому шарі і у верхніх частинах коріуму розташовані пігментні клітини. Шкіра забезпечує надійний захист від випаровування води, механічних пошкоджень і проникнення хвороботворних мікроорганізмів. Водночас вона втратила здатність до газообміну, випаровування води та виділення продуктів метаболізму. Шкіра щільно прилягає до тіла. Зміна рогового покриву проходить шляхом линьки: повного або часткового скидання старого рогового чохла і формування нового. Для багатьох видів линька проходить декілька разів на рік.
Пересування плазунів є різноманітним. При перебуванні в густих заростях трав, кронах дерев, в лісовій підстилці виявився зручним перехід до повзання особин, що призвело до втрати кінцівок. Особини пересуваються завдяки вигинанню тіла (змієподібний рух). Ця форма руху є ефективною також і у водному середовищі. Поява складок шкіри по боках тіла є передумовою для планерування тварин. Таким чином плазуни пересуваються різними способами (рис. 104).

Скелет. Хребетний стовбур плазунів ділиться на п’ять відділів: шийний, грудний, поперековий, крижовий і хвостовий.
При здобуванні їжі і орієнтуванні важливим для плазунів було формування гнучкої шиї і посилення рухливості голови. Рухливість голови забезпечується наявністю двох перших шийних хребців – атласа, або атланта (atlas), і епістрофея (epistropheus). Атлас має вигляд кісткового кільця, що розділений щільною зв’язкою на верхню і нижню половини; через верхній отвір головний мозок з’єднується з спинним; передня поверхня нижньої половини зчленовується з потиличним виростком черепа, а ззаду в нижній отвір входить зубоподібний відросток другого шийного хребця – епістрофея (рис. 107).
Рух шиї обумовлений числом і будовою хребців шийного відділу; вони є різними в різних групах. Гатерії мають хребці амфіцельні (риб’ячого типу) із залишками хорди між ними. У крокодилів і більшості лускатих хребці процельні (передньовгнуті) і лише у не багатьох нижчих форм амфіцельні. Частина шийних хребців несе короткі ребра. Складні рухи шиї черепах забезпечуються різноманіттям хребців: задні хребці процельні; передні – опістоцельні (задньовгнуті), а середні – амфіцельні.
До грудних хребців причленовуються довгі ребра, черевні кінці котрих з допомогою хрящових відділів прикріпляються до грудини, утворюючи закриту грудну клітку (немає грудної клітки у змій). До грудини причленовується плечовий пояс. Поперекові хребці також несуть ребра, котрі не доходять до грудини. До крижового відділу, який складаються із двох хребців, прикріпляється тазовий пояс. Хвостовий відділ допомагає зберегти рівновагу під час пересування, а інколи служить і рушієм руху (у водних змій, крокодилів і деяких водних ящірок). У ящірок, здатних до аутотомії, хвостові хребці можуть розламуватись посередині, тут є тонкі хрящові прошарки, що ділять тіло хребця на дві частини.
Загальне число хребців у різних видів плазунів може бути 50―80 (7―10 шийних, 16-25 грудинно-поперекових, 2 крижових, 15―40 хвостових), 140 (в товстих і коротких змій), 435 (у змій з довгим тілом).
П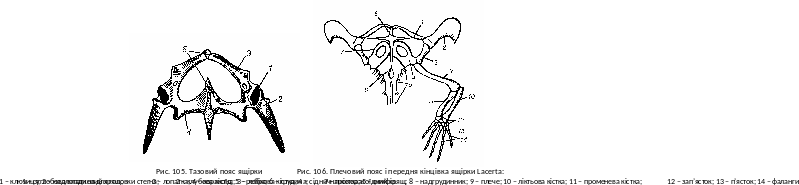 арні
кінцівки та їх пояси.
Плечовий пояс плазунів подібний до
поясу земноводних, але в ньому розвинуті
окостеніння коракоїд (coracoideum)
на місці з’єднанні з лопаткою (scapula)
утворюють суглобну ямку – місце для
приєднання головки плеча (рис. 106). Зверху
до лопатки приростає потовщений
надлопатковий хрящ (cartilago
suprascapularis),а
спереду до каракоїду – хрящовий
прокаракоїд (cart
procarocoidea).
Коракоїд і прокоракоїд кожної сторони
зростаються з непарною кістковою
грудиною (sternum);
через грудну клітку пояс передніх
кінцівок прикріпляється до осьового
скелету. Знизу до грудини приростає
хрестоподібна покривна кістка –
надгрудинник (episternum).
Парні покривні ключиці (clavicula)
з’єднують передній кінець нагдрудинника
з дорзальною частиною кожної лопатки.
арні
кінцівки та їх пояси.
Плечовий пояс плазунів подібний до
поясу земноводних, але в ньому розвинуті
окостеніння коракоїд (coracoideum)
на місці з’єднанні з лопаткою (scapula)
утворюють суглобну ямку – місце для
приєднання головки плеча (рис. 106). Зверху
до лопатки приростає потовщений
надлопатковий хрящ (cartilago
suprascapularis),а
спереду до каракоїду – хрящовий
прокаракоїд (cart
procarocoidea).
Коракоїд і прокоракоїд кожної сторони
зростаються з непарною кістковою
грудиною (sternum);
через грудну клітку пояс передніх
кінцівок прикріпляється до осьового
скелету. Знизу до грудини приростає
хрестоподібна покривна кістка –
надгрудинник (episternum).
Парні покривні ключиці (clavicula)
з’єднують передній кінець нагдрудинника
з дорзальною частиною кожної лопатки.
Т азовий
пояс складається із двох безіменних
кісток; кожна із них утворюється злиттям
трьох тазових кісток – клубової (ilium),
сідничної (ischium)
і лобкової (pubis).
Вони разом утворюють вертлужну ямку,
яка разом з головкою стегна формує
суглоб (рис. 105). Клубові кістки з’єднуються
з поперечними відростками крижових
хребців. У всіх сучасних плазунів таз
закритий: праві і ліві лобкові і сідничні
кістки одна з другою з’єднуються по
середній лінії симфізом – хрящовою
перемичкою.
азовий
пояс складається із двох безіменних
кісток; кожна із них утворюється злиттям
трьох тазових кісток – клубової (ilium),
сідничної (ischium)
і лобкової (pubis).
Вони разом утворюють вертлужну ямку,
яка разом з головкою стегна формує
суглоб (рис. 105). Клубові кістки з’єднуються
з поперечними відростками крижових
хребців. У всіх сучасних плазунів таз
закритий: праві і ліві лобкові і сідничні
кістки одна з другою з’єднуються по
середній лінії симфізом – хрящовою
перемичкою.
Парні кінцівки у різних видів і груп плазунів відрізняються в залежності від тих чи інших способів пересування. Але для них властива загальна схема будови парних кінцівок наземних хребетних (рис. 106).
Форма черепа плазунів залежить головним чином від характеру харчування та способів здобування їжі. Череп має витягнуті щелепи, які утворюють довге рило. Така форма щелеп дозволяє відривати куски тіла від великої здобичі. Одночасно зростає маса жувальної мускулатури. Череп майже повністю складається з кісток (рис. 107). Потиличний відділ формується із чотирьох кісток (оccіpitalіа) хондрального походження: верхньопотиличної; основної і двох бокових. Вони облямовують потиличний отвір, знизу якого розміщений єдиний потиличний відросток, утворений основною і двома боковими потиличними кістками. Покривна основна клиновидна кістка (basisphenoideum) лежить перед основною потиличною, і утворює дно черепа. Спереду до неї приростає невеликий парасфеноїд (parasphenoideum) і розташовуються парні леміші (voter), збоку від котрих лежать хоани. В ділянці слухової капсули виникають три вушні кістки (otiсi); передньовушна, є самостійною; задньовушна зростається із бічною потиличною, а верхньовушна, – зростається з верхньопотиличною. В нюховому відділі кісток немає, і він залишається хрящовим.
Д ах
черепа утворений парними покривними
кістками: носовими (nasalia), передлобними
(praefrоntalia);
лобними (frontalia) і задньолобними
(postfrontalia); дальше лежать тім’яні
(parietalia) і непарна міжтім’яна
(interparietalt)
кістки; остання має отвір для тім’яного
органу. Боки черепа утворюють покривні
кістки: парні міжщелепні (intermaxillare)
(у частини видів вони зливаються); парні
верхньощеплепні (maxillare);
надочні (supraorbitale);
виличні (jugale);
квадратно-виличні (gvadratojugale) і лускаті
(sgvamosum).
Піднебінно-квадратний хрящ в задній
своїй частині дає початок парним
хондральним окостенінням – квадратним
кісткам (qvadratum).
Передня частина піднебінно-квадратного
хряща замінюється покривними кістками,
які утворюють дно черепа: парними
піднебінними (palatini)
і криловидними (pterygoidei).
Поперечні кістки (transversi)
з’єднують криловидні кістки з
верхньощелепними, а в ящірок і гатерій,
крім того, верхньокриловидні, або
стовпчасті (epipterygoidei),
кістки зв’язують криловидні кістки
із тім’яними.
ах
черепа утворений парними покривними
кістками: носовими (nasalia), передлобними
(praefrоntalia);
лобними (frontalia) і задньолобними
(postfrontalia); дальше лежать тім’яні
(parietalia) і непарна міжтім’яна
(interparietalt)
кістки; остання має отвір для тім’яного
органу. Боки черепа утворюють покривні
кістки: парні міжщелепні (intermaxillare)
(у частини видів вони зливаються); парні
верхньощеплепні (maxillare);
надочні (supraorbitale);
виличні (jugale);
квадратно-виличні (gvadratojugale) і лускаті
(sgvamosum).
Піднебінно-квадратний хрящ в задній
своїй частині дає початок парним
хондральним окостенінням – квадратним
кісткам (qvadratum).
Передня частина піднебінно-квадратного
хряща замінюється покривними кістками,
які утворюють дно черепа: парними
піднебінними (palatini)
і криловидними (pterygoidei).
Поперечні кістки (transversi)
з’єднують криловидні кістки з
верхньощелепними, а в ящірок і гатерій,
крім того, верхньокриловидні, або
стовпчасті (epipterygoidei),
кістки зв’язують криловидні кістки
із тім’яними.
У черепах і в крокодилів розростання піднебінних відростків міжщелепних і верхньощелепних кісток, а також піднебінних кісток призводить до утворення вторинного кісткового піднебіння, яке розділяє ротову порожнину на верхній відділ – носоглотковий; і нижній – власне ротову порожнину.
Нижня щелепа утворюється із меккелового хряща, що замінюється хондрально-зчленівною кісткою (articulares), яка причленовується до квадратної, і низки шкірних кісток: зубної (dentale); кутової (anqulare), надкутової (supraangulare); вінцевої (coronare), а інколи ще декілька кісточок.
Верхній відділ під’язикової дуги (гіомандибуляре), перетворився у слухову кісточку середнього вуха – стремінце (stapes). Під’язиковй апарат складається із хрящової пластинки (гомолога копули) і трьох пар ріжків; гомологічних гіоїдам і залишкам зябрових дуг.
Еволюція жувальної мускулатури супроводжувалась перебудовою даху і частково бокових стінок первинно суцільно-стегального черепа, властивої предкам плазунів (рис. 108).
Змінилась також структура кісткової тканини. Загальний тип трубчастих кісток кінцівок зберігся, але товстоволокниста кісткова тканина властива земноводним змінилась на тонковолокнисту кістку, яка є більш міцнішою. При цьому кістки кінцівок набули остеонну структуру і зонарну (шарову) будову. Кістковий елемент – остеон – представляє собою кісткову трубку, стінки котрої утворені тонкими, але міцними кістковими пластинками, а порожнина кісткової тканини зайнята кровоносною судиною і нервом.
Мускулатура плазунів, у порівнянні з амфібіями, суттєво змінилася. Вихідне метамерне розташування мускулатури, збереглося лише в розміщенні м’язів, що з’єднують поряд розташовані хребці, і в мускулатурі черевної стінки. На черепі у скроневих ямках розвивається могутня жувальна мускулатура, яка приводить в рух щелепи. Поява рухомого шийного відділу супроводжувалась розвитком шийної мускулатури, особливо добре розвиненої у особин видів з видовженими й гнучкішими шиями. Стрічкоподібні м’язи мускулатури беруть участь у згинанні тіла та русі кінцівок.. Появляються зачатки підшкірної мускулатури, яка змінює положення рогових лусок. Така мускулатура добре розвинена у змій та безногих ящірок і бере участь в переміщенні тіла, забезпечуючи координовані рухи черевних щитків.
У зв’язку з переходом плазунів на легеневе дихання, покращилась вентиляція легень. Появляється грудна клітка, яку в рух приводить спеціальна міжреберна мускулатура. Зміцнилися м’язи черевного пресу.
Органи травлення та живлення. Сучасні плазуни є переважно тваринноїдними видами. Більшість харчуються дрібними наземними і водними тваринами, запаси котрих в природі великі і більш-менш сталі; поїдають також дрібних хребетних – риб, земноводних, мишовидних гризунів та ін. Водні і навколоводні черепахи та змії харчуються водними безхребетними й рибою. Переважно головоногими молюсками живляться морські змії, наприклад, Pelamis platurus. Крокодили і великі змії нападають на крупну здобич. Наземні і водні черепахи, тропічні агами й ігуани є рослинноїдними плазунами.
Захоплення їжі рептилії проводять за допомогою щелеп, які озброєнні численними гострими зубами. Зуби приростають до щелеп і піднебінних кісток (плевродонтні і акродонтні зуби); лише у крокодилів і викопних звірозубих плазунів основа зубів занурена в спеціальні комірки – альвеоли (текодонтні зуби). У сучасних плазунів зуби однотипні; лише у частини змій розвиваються спеціалізовані отруйні зуби. Крокодили і черепахи здатні відривати від крупної здобичі окремі куски. Більшість видів проковтують їжу повністю. Черепахи, в яких немає зубів зрізають рослини гострими краями рогового покриву щелеп. Будова щелепного апарату змій дозволяє широко відкривати пащу (рис. 109) і проковтувати здобич, яка перевищує діаметр змії. Тому в черепі змій зникли скроневі дуги, а щелепний апарат перетворився на шарнірно-важилеву систему.
В ротовій порожнині рептилій розташовані
слинні залози, слизовий секрет котрих
змочує їжу і полегшує її заковтування.
В отруйних змій і окремих видів ящірок
деякі слинні залози перетворилися в
отруйні, секрет котрих стікає каналом
або поверхневою борозною отруйних
зубів. Частина залоз набула здатність
виробляти травні ферменти (вміст їх в
слині плазунів незначний).
ротовій порожнині рептилій розташовані
слинні залози, слизовий секрет котрих
змочує їжу і полегшує її заковтування.
В отруйних змій і окремих видів ящірок
деякі слинні залози перетворилися в
отруйні, секрет котрих стікає каналом
або поверхневою борозною отруйних
зубів. Частина залоз набула здатність
виробляти травні ферменти (вміст їх в
слині плазунів незначний).
Н а
дні ротової порожнини розташований
мускулистий язик, який тварина здатна
далеко висувати. Ящірки та змії мають
роздвоєний язик, що використовується
як орган нюху, а разом з якобсоновим
органом – і як хеморецептор. Розширений
на кінці язик хамелеонів може раптово
викидатися і служить для захоплення
дрібної рухомої здобичі (комахи та ін.)
(рис. 110).
а
дні ротової порожнини розташований
мускулистий язик, який тварина здатна
далеко висувати. Ящірки та змії мають
роздвоєний язик, що використовується
як орган нюху, а разом з якобсоновим
органом – і як хеморецептор. Розширений
на кінці язик хамелеонів може раптово
викидатися і служить для захоплення
дрібної рухомої здобичі (комахи та ін.)
(рис. 110).
Д обре
виражений стравохід. У змій він має
міцну мускулатуру, яка проштовхує
крупну здобич до шлунка. Відмежований
від стравоходу шлунок має мускулисті
стінки. Кишечник довший, ніж у земноводних.
На межі між тонкою і товстою кишкою
відходить первинна сліпа кишка; вона
краще розвинена у рослиноїдних (степова
черепаха та ін.). Кишечник відкривається
в клоаку (рис. 111). Підшлункова залоза
лежить у першій петлі кишечника. Крупна
печінка має жовчний міхур, протока
якого відкривається в кишечник поруч
з підшлунковою залозою.
обре
виражений стравохід. У змій він має
міцну мускулатуру, яка проштовхує
крупну здобич до шлунка. Відмежований
від стравоходу шлунок має мускулисті
стінки. Кишечник довший, ніж у земноводних.
На межі між тонкою і товстою кишкою
відходить первинна сліпа кишка; вона
краще розвинена у рослиноїдних (степова
черепаха та ін.). Кишечник відкривається
в клоаку (рис. 111). Підшлункова залоза
лежить у першій петлі кишечника. Крупна
печінка має жовчний міхур, протока
якого відкривається в кишечник поруч
з підшлунковою залозою.
Перетравлення плазунами великої здобичі проходить нормально лише при досить високій температурі середовища; затримка перетравлення при низьких температурах може зумовити отруєння і загибель тварини. Плазуни (черепахи, змії) здатні до тривалого голодування. Деякі змії й черепахи можуть жити у неволі один-два роки без їжі. Ящірки в активному стані можуть обходитися без їжі багато тижнів.
О ргани
дихання і газообмін.
Зародок, що розвивається в яйці плазунів,
онтогенестично відповідає стадії
личинки земноводних, дихає за допомогою
кровоносних капілярів жовткового
мішка, а згодом – алантоїса. Шкіра
плазунів покрита роговими утворами і
не бере участі в диханні. Основними
органами дихання після вилуплення із
яйця служать парні легені; у змій права
легеня більша (у амфібій – ліва). Легені
мають мішкоподібну будову, але їхня
внутрішня структура набагато складніша,
ніж у земноводних (рис. 112). Вентиляція
легень забезпечується роботою грудної
клітки з допомогою міжреберної і
черевної мускулатури. Складна будова
легень у черепах, які здатні засвоювати
кисень навіть при слабій вентиляції
легень, пов’язана з утворенням панцира.
ргани
дихання і газообмін.
Зародок, що розвивається в яйці плазунів,
онтогенестично відповідає стадії
личинки земноводних, дихає за допомогою
кровоносних капілярів жовткового
мішка, а згодом – алантоїса. Шкіра
плазунів покрита роговими утворами і
не бере участі в диханні. Основними
органами дихання після вилуплення із
яйця служать парні легені; у змій права
легеня більша (у амфібій – ліва). Легені
мають мішкоподібну будову, але їхня
внутрішня структура набагато складніша,
ніж у земноводних (рис. 112). Вентиляція
легень забезпечується роботою грудної
клітки з допомогою міжреберної і
черевної мускулатури. Складна будова
легень у черепах, які здатні засвоювати
кисень навіть при слабій вентиляції
легень, пов’язана з утворенням панцира.
Новий спосіб дихання супроводжується перебудовою дихальних (повітроносних) шляхів: утворюється не спадаюча дихальна трубка – трахея, стінки котрої підтримуються еластичними хрящовими кільцями. Вхід до трахеї (із гортанної камери) облямований перстнеподібним і парними черпалоподібними хрящами; в ротову порожнину камера відкривається гортанною щілиною. На задньому кінці трахея ділиться на два бронхи, що впадають в легені й галузяться там на дрібніші трубки; стінки бронхів також зміцнені кільцями. Ритм дихання змінюється, залежно від величини зовнішньої температури і стану тварини. Так, у ящірки Sceloporus частота дихання при 15оС становила 26 дихальних рухів на хвилину, а при 25―35оС – уже 37.
Кровоносна система і кровообіг. У плазунів серце трикамерне. Передсердя розділене повною перегородкою; кожне відкривається в шлуночок самостійним отвором, який містить клапан із півмісяцевих складок. Шлунок має неповну перегородку, яка розділяє його на дві частини: в період систоли перегородка доходить до спинної стінки шлуночка, на короткий час, повністю розділяє його, що призводить до розділення потоків крові з різним вмістом кисню. Венозна пазуха злита з правим передсердям.
А ртеріальний
конус редукований, і від різних ділянок
шлуночка відходять самостійно три
судини. Від правої частини шлуночка,
що вміщує венозну кров, відходить
легенева артерія, яка ділиться на праву
й ліву. Від лівої частини шлуночка, що
вміщує артеріальну кров, починається
права дуга аорти, від якої відділяються
сонні й підключичні артерії. Від середини
шлуночка відходить ліва дуга аорти
(рис. 113). Обігнувши серце, ліва і права
дуги аорти зливаються у спинну аорту.
До легеневої артерії надходить венозна
кров, а в праву дугу аорти і у сонні і
підключичні артерії, що відходять від
неї – артеріальна. В ліву дугу аорти
проходить змішана кров. Тому по спинній
аорті тече змішана кров, з перевагою
артеріальної.
ртеріальний
конус редукований, і від різних ділянок
шлуночка відходять самостійно три
судини. Від правої частини шлуночка,
що вміщує венозну кров, відходить
легенева артерія, яка ділиться на праву
й ліву. Від лівої частини шлуночка, що
вміщує артеріальну кров, починається
права дуга аорти, від якої відділяються
сонні й підключичні артерії. Від середини
шлуночка відходить ліва дуга аорти
(рис. 113). Обігнувши серце, ліва і права
дуги аорти зливаються у спинну аорту.
До легеневої артерії надходить венозна
кров, а в праву дугу аорти і у сонні і
підключичні артерії, що відходять від
неї – артеріальна. В ліву дугу аорти
проходить змішана кров. Тому по спинній
аорті тече змішана кров, з перевагою
артеріальної.
В
Рис. 113. Схема кровоносної системи ящірки (за Огньовим зі змінами):
1 – легенева артерія; 2 – права дуга аорти; 3 – сонна артерія; 4 – сонний протік; 5 – підключична артерія; 6 – ліва дуга аорти; 7 – спинна аорта; 8 – міжхребтова артерія; 9 – кишкова артерія; 10 – брижова артерія; 11 – клубова артерія; 12 – стегнова артерія; 13 – печінка; 14 – нирка; 15 – хвостова артерія; 16 – хвостова вена; 17 – стегнова вена; 18 – порожниста вена; 19 – черевна вена; 20 – ниркова вена; 21 – задня порожниста вена; 22 – ворітня вена печінки; 23 – яремна вена; 24 – підключична вена; 25 – 26 – ліва і права передні порожнисті вени; 27 – відрізок травної трубки; 28 – легеня.
енозна система плазунів подібна до венозної системи земноводних. Хвостова вена (рис. 113) в ділянці таза ділиться на дві клубові або тазові вени, до яких підходять вени від задніх кінцівок. Клубові вени відділяють від себе ворітні вени нирок і зливаються в черевну вену. Черевна вена, разом з ворітною веною печінки, що несе кров від кишечника, розпадається в печінці на капіляри. В печінці проходить детоксикація продуктів розпаду білків, починається синтез продуктів виділення, відкладаються запаси глікогену і здійснюються деякі процеси кровотворення. Капіляри ворітної системи печінки зливаються в печінкові вени, які впадають в задню порожнисту вену. Від голови кров несуть парні яремні вени. Вони з’єднуються з підключичними венами, утворюють праву і ліву передні порожнисті вени, які впадають в праве передсердя. У ліве передсердя входить легенева вена, яка утворилась внаслідок злиття правої і лівої легеневих вен (несуть артеріальну кров).Кількість крові та вміст гемоглобіну в організмі плазунів лише дещо більший, ніж у земноводних.
Органи виділення і водно-сольовий обмін. Серед пристосувань, що дозволили плазунам перейти до наземного способу життя, важливим є заміна мезонефричної (тулубової) нирки (круглороті, риби, земноводні) метанефричною (тазовою) і пов’язана з цим перебудова водно-сольового обміну. При цьому змінився склад виведених із організму продуктів азотистого обміну. Його кінцевими продуктами є декілька речовин – аміак, сечова кислота, сечовина, креатин, креатинін та ін. Метанефрична тазова нирка відрізняється не лише положенням (розташована в тазовій ділянці) а, й мікроструктурою. Вона має спрощений гломерулярний апарат і ускладнену будову ниркових (нефронних) канальців. Клубочки метанефросу мають дві-три капілярні петлі, тому володіють меншою здатністю щодо фільтрації. У зв’язку з цим зростає значення ниркових канальців, де не лише йде реабсорбція води і солей натрію, але й виділення їх секреторними клітинами сечовини і сечової кислоти. В результаті концентрація цих речовин на виході із канальця стає у 20 раз вищою, ніж їх концентрація в крові. Канальці видовжені й діляться на зігнутий проксимальний, проміжний, зігнутий дистальний і збірний відділи. В перших трьох відділах проходять секреторні процеси і обернене всмоктування води і цінних для організму, низькомолекулярних сполук (цукрів, вітамінів, амінокислот і ін.), останній відділ має простий канал, через який виводить секрет. Збагачена продуктами виділення кінцева сеча стікає з нирок через сечопроводи у клоаку й сечовий міхур, де завершується поглинання води, після чого максимально концентрована сеча виводиться із організму. Інтенсивність фільтрації й реабсорбція речовин в канальцях, клоаці й сечовому міхурі залежить від ступеня проникності їхніх стінок. Остання регулюється антидіуретичними й деякими іншими гіпофізарними гормонами.
Тазові нирки розвиваються із зачатків, що утворюються, ззаду тулубових нирок, зародків, які в подальшому редукуються. Метанефричні нирки у рептилій мають вигляд двох компактних тіл, розташованих біля спинної стінки тазової ділянки. В ході їхнього розвитку від задньої частини вольфових каналів (мезонефричних проток) відшнуровується пара самостійних сечопроводів. Вони впадають у клоаку зі спинного боку. У видів, у виділенні котрих переважає сечовинай інші розчинні речовини, добре розвинений сечовий міхур, що відкривається в клоаку з черевного сбоку. Види, в сечі котрих переважає сечова кислота, мають слабше розвинений сечовий міхур (крокодили, змії, деякі ящірки). Морські черепахи і деякі інші плазуни, які вимушені використовувати солену воду для пиття, мають особливі залози, що виводять надлишок солей із організму. У черепах вони розташовані в очній ямці. Секрет сольових залоз морських плазунів вміщує семипроцентний розчин кухонної солі, тому вони дійсно «плачуть гіркими сльозами», звільняючись від надлишку солей. Морські ігуани, які харчуються насиченими сіллю водоростями, мають сольові залози у вигляді так званих «носових залоз», протоки котрих відкриваються в носову порожнину. Солі виділяються у вигляді крапель, що витікають із носових отворів.
Статеві органи і розмноження. Статеві залози лежать в порожнині тіла з боків хребта. Сім’яники – це парні овальні тіла, які з’єднані з сім’япроводами. Правий і лівий сім’япровід відкриваються у відповідні сечопроводи. Самці всіх плазунів, крім гатерії, мають спеціальні копулятивні органи; у крокодилів і черепах непарний, а у ящірок та змій – парні вирости задньої стінки клоаки, котрі під час запліднення вивертаються назовні (рис. 114).
П арні
яєчники мають вигляд зернистих овальних
тіл. Від них відходять мюлерові канали.
Вони починаються миготливими лійками,
розташованими біля яєчників, а
закінчуються отворами в клоаці.
Запліднення проходить у верхньому
відділі яйцепроводу. Виділення
секреторних залоз середньої частини
яйцепроводу утворює довкола яйцеклітини
(жовтка) білкову оболонку. Вона погано
розвинена у змій і ящірок, і добре – у
черепах та крокодилів. Із секрету, що
виділяють клітини стінок нижньої
частини яйцепроводу (матки), формуються
зовнішні оболонки яйця.
арні
яєчники мають вигляд зернистих овальних
тіл. Від них відходять мюлерові канали.
Вони починаються миготливими лійками,
розташованими біля яєчників, а
закінчуються отворами в клоаці.
Запліднення проходить у верхньому
відділі яйцепроводу. Виділення
секреторних залоз середньої частини
яйцепроводу утворює довкола яйцеклітини
(жовтка) білкову оболонку. Вона погано
розвинена у змій і ящірок, і добре – у
черепах та крокодилів. Із секрету, що
виділяють клітини стінок нижньої
частини яйцепроводу (матки), формуються
зовнішні оболонки яйця.
Більшість плазунів закопують відкладені яйця в ґрунт у добре прогрітих місцях; частина видів відкладає яйця в купи рослинного сміття або під гнилі ппеньки, використовуючи тепло, що утворюється під час гниття. Деякі крокодили риють ями й засипають яйця рослинними рештками; самки тримаються біля гнізда і охороняють кладку. Деякі дрібні ящірки також охороняють свої кладки (варани та ін.). Самки пітонів обкручують кладку яєць своїм тілом, охороняють і обігрівають її. В такому «гнізді» температура на 6―12 оС вища, ніж в оточуючому середовищі. У крокодилів самка розкопує кладку після вилуплення малят, полегшуючи їм вихід на поверхню; самки частини видів охороняють дітей і на початку їхнього самостійного життя.
Для невеликої кількості видів сучасних лускатих властиве яйцеживородіння або, зрідка, живородіння (гадюка звичайна, веретільниця).
Нервова система й органи чуття, нервова діяльність. Головний мозок плазунів відрізняється від головного мозку земноводних, зокрема передній мозок крупніший; його збільшення зв’язане з розвитком мозкового склепіння півкуль та збільшенням смугастих тіл (corpora striata), які й складають більшу частину маси переднього мозку (рис. 115). В мозковому склепінні чітко помітно первинне склепіння, або архіпаліум, яке займає велику частину покрівлі півкуль, а також зачаток неопаліума. Нюховий центр плазунів, у порівнянні з амфібіями, більш складний і диференційований. Проміжний мозок згори прикритий півкулями переднього мозку. В його покрівлі розташовані епіфіз (ендокринна залоза) і тім’яний орган, який здатний сприймати світлові сигнали.
Зорова кора середнього мозку більш розвинена, ніж у амфібій. Мозочок плазунів крупний, що відповідає значному ускладненню й інтенсивності їхнього руху. Продовгуватий мозок утворює згин у вертикальній площині, що характерний для всіх амніот. Він координує основні вегетативні функції (дихання, кровообіг, травлення та ін.). Від головного мозку відходить 11 пар головних нервів.
В спинному мозку чітко помітний поділ білої (провідних шляхів) і сірої (нервових клітин) речовини. Це свідчить про посилення контролю центрів головного мозку над рефлекторними механізмами спинного мозку.
Орган зору пристосований до роботи у повітряному середовищі. Око захищене зовнішніми повіками і миготливою перетинкою. Змії та деякі ящірки (гекони, сцинки, частина безногих ящірок) мають зрощені повіки, які творять прозору оболонку. У нічних видів очі збільшені, мають вертикальну зіницю. Сльозові залози захищають око від висихання. Акомодація ока досягається переміщенням кришталика і зміною його кривизни за допомогою поперечносмугастого війчастого м’яза. Відчуття кольорового зору для більшості плазунів зміщене в жовто-оранжеву частину спектра. На відміну від земноводних, аналіз і синтез зорових образів відбувається не на сітківці, а, переважно, у зоровій півкулі середнього мозку.
У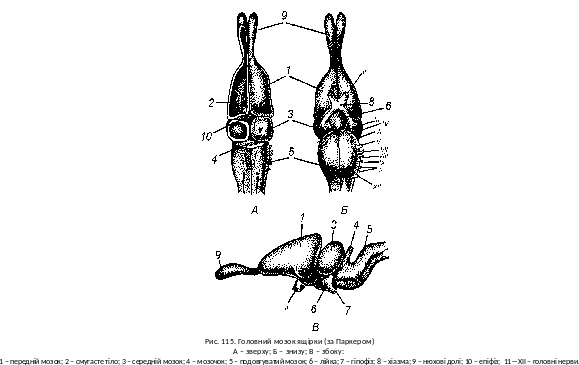 просторовій орієнтації і спілкуванні
плазунів зір відіграє важливу роль.
Ямкоголові змії (Crotalidae),
пітони (Pythoninae)
і африканські гадюки (Bitis)
мають особливі органи термічного
відчуття – терморецептори, і, навіть,
термолокатори. Термолокатори ямкоголових
змій – парні ямки, які розташовані з
боків морди, між ніздрями й очима, у
пітонів подібні неглибокі ямки є на
верхньогубних щитках.
просторовій орієнтації і спілкуванні
плазунів зір відіграє важливу роль.
Ямкоголові змії (Crotalidae),
пітони (Pythoninae)
і африканські гадюки (Bitis)
мають особливі органи термічного
відчуття – терморецептори, і, навіть,
термолокатори. Термолокатори ямкоголових
змій – парні ямки, які розташовані з
боків морди, між ніздрями й очима, у
пітонів подібні неглибокі ямки є на
верхньогубних щитках.
Орган слуху плазунів подібний до органу слуху жаб. Складається із середнього вуха з барабанною перетинкою і однією слуховою кісточкою – стремінцем, яке передає коливання перетинки на кругле віконце, що відділяє порожнину внутрішнього вуха. У внутрішньому вусі відокремлюється завиток (равлик Ладена), який служить апаратом аналізу і кодування акустичних сигналів. Він має просту будову і у більшості видів представляє мішкоподібний виріст. Плазуни сприймають звуки в діапазоні 20―6000 Гц, але більшість добре чує в діапазоні 60―200 Гц. Крокодили краще сприймають звуки частотою 100―3000 Гц. Змії мають поганий слух; вони позбавлені барабанної перетинки, сприймають, переважно, звуки, які поширюються в грунті або воді (так званий сейсмічний слух). Так само сприймають звук і змієподібні ящірки.
Більшість плазунів безголосі. Звуки змій (шипіння, хрип, стук хвостових брязкалець) і деяких ящірок (скрип лусок) частіше мають значення попередження. Голосні ревучі звуки властиві крокодилам; під час охорони території або пошуку особин іншої статті.
Важливу роль в орієнтації і в спілкуванні рептилій відіграють хеморецептори. Органи нюху відкриваються назовні парними ніздрями, а в порожнину рота – щілиноподібними хоанами. Багато ящірок за допомогою нюху знаходять їжу, викопують її з піску з глибини до 6―8 см. Варани, полози і гадюки за допомогою нюху можуть розрізняти особин свого або чужих видів. Черепахи, ящірки, крокодили мають спеціальні пахучі залози, секретом котрих мітять зайняту територію, попереджаючи вторгнення на неї інших особин.
Сучасна фауна плазунів нараховує біля 300 видів. Нижче подаємо їх огляд.
КЛАСИФІКАЦІЯ ПЛАЗУНІВ