

Клас земноводні, або амфібії – amphibia
Земноводні, або амфібії є першими наземними хребетними, які однак зберегли тісний зв’язок з водним середовищем. У більшості видів яйця (ікра) позбавлені щільних оболонок і можуть розвиватися лише у воді. Личинки, які вилупились із яєць, ведуть водний спосіб життя і згодом проходять метаморфоз (перетворення), під час якого формуються ознаки дорослих особин, пристосованих до життя на суходолі.
Дорослі земноводні мають парні кінцівки з шарнірними суглобами. Череп з допомогою двох потиличних виростків рухомо з’єднується з шийним хребцем. Піднебінно-квадратний хрящ зростається з мозковою коробкою (аутостилічний череп), а верхній елемент під’язикової дуги – підвісок – перетворюється в кісточку середнього вуха – стремінце. Тазовий пояс причленовується до поперечних відростків крижового хребця. Формуються два кола кровообігу, які повністю не роз’єднані: серце має два передсердя, але шлуночок один. Очі закриваються рухомими повіками. Органи бічної лінії в дорослих тварин зникають. Передній мозок збільшується і розділяється на дві півкулі, зверху яких нагромаджуються нервові клітини. У амфібій рівень метаболізму дещо вищий, ніж у риб. Шкіра без будь-яких кісткових або хрящових включень, містить велику кількість залоз, які виділяють слиз. Через шкіру відбувається проникнення води і газів. Органами виділення служать тулубові (мезонефричні) нирки та шкіра. Температура тіла земноводних залежить від температури середовища (пойкілотермія).
Зараз налічують понад 2500 видів земноводних.
Особливості організації земноводних
Форма тіла. Сучасні земноводні мають короткий, сплющений дорзовентрально тулуб з редукованим хвостом, задні кінцівки довші і міцніші від передніх (ряд безхвості); або ж тіло валькувате, видовжене, інколи трохи сплюснуте, або стиснуте з боків, з невеликою головою, довгим хвостом і короткими кінцівками (ряд хвостаті); нарешті, серед представників класу трапляються тварини, позбавлені кінцівок, червоподібні, з маленькою головою (ряд безногі). Розміри земноводних коливаються: безхвості мають довжину 3―25 см.; хвостаті 10―30 см і окремі великі (гігантська саламандра до 1,6 м); безногі (червуги) досягають 30-120 см в довжину.
П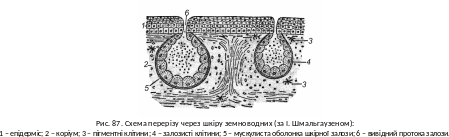 окриви,
шкіра та їх похідні.
Епідерміс багатошаровий, коріум тонкий,
але сильно насичений капілярами. Шкіра
у земноводних має численні багатоклітинні
залози (рис. 87). Вони виділяють тонкий
шар слизу, що покриває все тіло, зволожуючи
шкіру і тим самим охороняє її від
висихання. Шкіра бере участь в газообміні.
У ропух, які живуть у відносно сухих
місцях слизь гусне і утворює на шкірі
щільну плівку, завдяки якій знижується
віддача вологи. Секрет шкірних залоз
(джерлянок, ропух, деяких саламандр) має
подразливі або отруйні речовини. В
секреті містяться також речовини, які
мають сигнальне значення, завдяки чому
вони впливають на поведінку інших
особин. В нижніх шарах епідермісу та в
коріумі розташовані пігментні клітини,
які зумовлюють видоспецифічне забарвлення.
Останнє виконує різні функції: маскування
(криптичне або захисне забарвлення);
попередження та відлякування для видів
з отруйними залозами (апосематичне
забарвлення з яскравими кольоровими
плямами); статевих відмін – у самців
забарвлення досить часто стає більш
яскравим на початку розмноження, що
полегшує зустріч статевозрілих особин
і стимулює спарювання. Окремі види
можуть змінювати інтенсивність
забарвлення в залежності від кольору
фону, найкраще це виражено в деяких
деревних жаб.
окриви,
шкіра та їх похідні.
Епідерміс багатошаровий, коріум тонкий,
але сильно насичений капілярами. Шкіра
у земноводних має численні багатоклітинні
залози (рис. 87). Вони виділяють тонкий
шар слизу, що покриває все тіло, зволожуючи
шкіру і тим самим охороняє її від
висихання. Шкіра бере участь в газообміні.
У ропух, які живуть у відносно сухих
місцях слизь гусне і утворює на шкірі
щільну плівку, завдяки якій знижується
віддача вологи. Секрет шкірних залоз
(джерлянок, ропух, деяких саламандр) має
подразливі або отруйні речовини. В
секреті містяться також речовини, які
мають сигнальне значення, завдяки чому
вони впливають на поведінку інших
особин. В нижніх шарах епідермісу та в
коріумі розташовані пігментні клітини,
які зумовлюють видоспецифічне забарвлення.
Останнє виконує різні функції: маскування
(криптичне або захисне забарвлення);
попередження та відлякування для видів
з отруйними залозами (апосематичне
забарвлення з яскравими кольоровими
плямами); статевих відмін – у самців
забарвлення досить часто стає більш
яскравим на початку розмноження, що
полегшує зустріч статевозрілих особин
і стимулює спарювання. Окремі види
можуть змінювати інтенсивність
забарвлення в залежності від кольору
фону, найкраще це виражено в деяких
деревних жаб.
В наземних видів плоскі клітини зовнішнього шару епітелію піддаються зроговінню на кінцях пальців, утворюючи кігті (шпорцева жаба, а також в Примор’ї трапляється кігтистий тритон – Onychodactylus fischeri). У безногих в коріумі містяться дрібні кісткові луски – залишки кісткового покриву палеозойських земноводних. В безхвостих земноводних під шкірою розташовані великі лімфатичні лакуни (резервуари), в яких при сприятливих умовах накопичується запас води.
Рухова система та основні типи рухів. Характер руху земноводних є досить одноманітним. Розрізняють два основні типи. Викопні і сучасні хвостаті земноводні зберегли тип руху притаманний рибам – за допомогою сильних бокових згинів всього тіла, але з опорою на короткі ноги під час руху по землі. При коротких кінцівках бокові згини тулуба збільшують довжину кроку, а згини хвоста допомагають зберігати рівновагу. У воді кінцівки не виконують подібної ролі. Безногі пересуваються з допомогою згинів всього тіла.
Б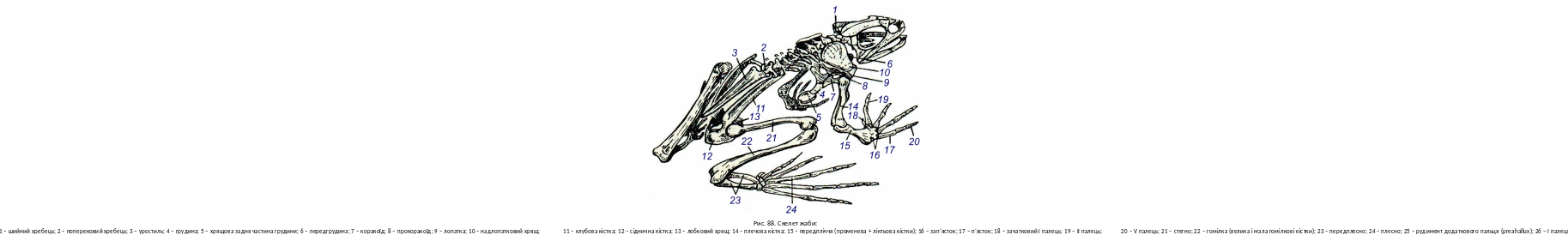 езхвості
земноводні на суші пересуваються
стрибками, піднімаючи тіло у повітря
різким поштовхом обох задніх кінцівок.
Коротконогі види (ропухи) крім стрибків
можуть повільно крокувати, послідовно
переставляючи кінцівки. У воді, безхвості
енергійно плавають, за допомогою задніх
кінцівок (стиль «брас», але без участі
передніх кінцівок). Рух стрибками призвів
до вкорочення і дорзовентрального
сплющення тулуба, зникнення хвоста,
видовження задніх кінцівок, вироблення
ряду специфічних рис в будові скелета
(зменшення числа хребців, їх міцному
з’єднанні, видовженню клубових кісток
та ін.).
езхвості
земноводні на суші пересуваються
стрибками, піднімаючи тіло у повітря
різким поштовхом обох задніх кінцівок.
Коротконогі види (ропухи) крім стрибків
можуть повільно крокувати, послідовно
переставляючи кінцівки. У воді, безхвості
енергійно плавають, за допомогою задніх
кінцівок (стиль «брас», але без участі
передніх кінцівок). Рух стрибками призвів
до вкорочення і дорзовентрального
сплющення тулуба, зникнення хвоста,
видовження задніх кінцівок, вироблення
ряду специфічних рис в будові скелета
(зменшення числа хребців, їх міцному
з’єднанні, видовженню клубових кісток
та ін.).
Скелет земноводних складається з хребта або осьового стовбура, черепа, кінцівок та їх поясів (малого тазу). Хребет ділиться на 4 відділи: шийний, тулубовий, поперековий (чи крижовий) і хвостовий (рис. 88). Шийний і поперековий відділи мають по одному хребцю. Перший забезпечує деяку рухливість голови щодо тулуба, поперековий слугує для зчленування з тазовим поясом. Тулубових хребців у безхвостих амфібій – 7; всі хвостові хребці (їх 12) зливаються в єдину кістку – уростиль. Хвостаті мають 13―62 тулубових і 22―36 хвостових хребців; загальне число хребців у безногих сягає 200―300.
У примітивних земноводних (безногих частково хвостатих, гладконогих і безхвостих) хребці, як і в риб, амфіцельні; між тілами хребців та в них зберігаються залишки хорди. У справжніх саламандр і частини безхвостих (круглоязикові і піпові) хребці опістоцельні (тіла хребців спереду випуклі, ззаду вгнуті) у решти безхвостих – процельні (спереду вгнуті, ззаду випуклі). Над тілами хребців добре виражені верхні дуги, які утворюють канал для спинного мозку. Тулубові хребці мають добре розвинуті поперечні відростки, до яких у хвостатих причленовуються дуже короткі ребра. Поперечні відростки шийного хребця розвинуті слабо. Хвостові хребці хвостатих несуть нижні дуги, утворюючи, (як і в риб) гемальний канал.
Ч ереп
земноводних у дорослому стані містить
багато хряща. В порівнянні з костистими
рибами в сучасних земноводних кісток
менше, тоді як у древніх вимерлих видів
покривних кісток було більше. В потиличному
відділі осьового черепа розвиваються
парні бокові потиличні кістки (occipitale
laterale)
(рис. 89) що мають потиличні виростки. Два
потиличні виростки зчленовуються з
шийним хребцем, – характерна риса
амфібій і ссавців, у рептилій і птахів
є один виросток. В слуховому відділі є
одна пара кісток – передньовушна
(prooticum).
В очному відділі хвостатих амфібій
лежать парні очно-клиноподібні кістки
(orbitosphenoideum);
у безхвостих вони зливаються в одну
кільцеподібну клиноподібно-нюхову
кістку (sphenethmoideum).
Решта частина мозкової коробки залишається
хрящовою. Незначне число і покривних
кісток. Покрівлю черепа утворюють парні
тім’яні кістки, у безхвостих вони
зливаються в парні лобно-тім’яні кістки
(frontoparietale).
Спереду лежать парні носові кістки
(nasale),
а у хвостатих ще 1―2 пари передлобних
кісток (praefrontale).
В слуховому відділі формується покривна
луската кістка (squamosum).
Дно черепа вкриває крупний парасфеноїд
(parasphenoideum),
спереду якого лежать парні покривні
кістки піднебінні (palatinum)
і леміші (vomer);
у хвостатих вони зливаються в парні
піднебінно-лемішеві кістки (vomeropalatinum).
На лемішах, а у хвостатих і на піднебінних
кістках сидять дрібні зуби.
ереп
земноводних у дорослому стані містить
багато хряща. В порівнянні з костистими
рибами в сучасних земноводних кісток
менше, тоді як у древніх вимерлих видів
покривних кісток було більше. В потиличному
відділі осьового черепа розвиваються
парні бокові потиличні кістки (occipitale
laterale)
(рис. 89) що мають потиличні виростки. Два
потиличні виростки зчленовуються з
шийним хребцем, – характерна риса
амфібій і ссавців, у рептилій і птахів
є один виросток. В слуховому відділі є
одна пара кісток – передньовушна
(prooticum).
В очному відділі хвостатих амфібій
лежать парні очно-клиноподібні кістки
(orbitosphenoideum);
у безхвостих вони зливаються в одну
кільцеподібну клиноподібно-нюхову
кістку (sphenethmoideum).
Решта частина мозкової коробки залишається
хрящовою. Незначне число і покривних
кісток. Покрівлю черепа утворюють парні
тім’яні кістки, у безхвостих вони
зливаються в парні лобно-тім’яні кістки
(frontoparietale).
Спереду лежать парні носові кістки
(nasale),
а у хвостатих ще 1―2 пари передлобних
кісток (praefrontale).
В слуховому відділі формується покривна
луската кістка (squamosum).
Дно черепа вкриває крупний парасфеноїд
(parasphenoideum),
спереду якого лежать парні покривні
кістки піднебінні (palatinum)
і леміші (vomer);
у хвостатих вони зливаються в парні
піднебінно-лемішеві кістки (vomeropalatinum).
На лемішах, а у хвостатих і на піднебінних
кістках сидять дрібні зуби.
У вісцеральному відділі черепа піднебінно-квадратний хрящ зберігається протягом всього життя. Переднім і заднім кінцями він приростає до черепної коробки (аутостилія) (рис. 89). До піднебінно-квадратного хряща прилягають парні покривні кістки – передщелепова (praemaxillare) і верхньощелепова (maxillare). На згаданих щелепових кістках сидять дрібні зуби; у частини видів, наприклад, жаб, вони редукуються. Задня ділянка піднебінно-квадратного хряща зверху вкрита покривною квадратно-виличною кісткою (quadrato-jugale) та лускатою кісткою, а знизу – крилоподібною кісткою (pterygoideum). У деяких хвостатих земноводних задня ділянка піднебінно-квадратного хряща костеніє, утворюючи маленьку квадратну кістку (quadratum). Первинна нижня щелепа – меккелів хрящ – залишається хрящовим і лише її передній кінець костеніє в маленькі парні підборіддєво-щелепові кістки (mento-mandibulare). За ними, прикриваючи меккелів хрящ, лежать покривні зубні кістки (dentale), які у сучасних земноводних позбавлені зубів. Задню частину меккелевого хряща обростає видовжена покривна кутова кістка (angulare) і кілька дрібних додаткових покривних кісточок. Суглобним відростком меккелів хрящ з’єднується з заднім кінцем піднебінно-квадратного хряща, утворюючи щелеповий суглоб.
П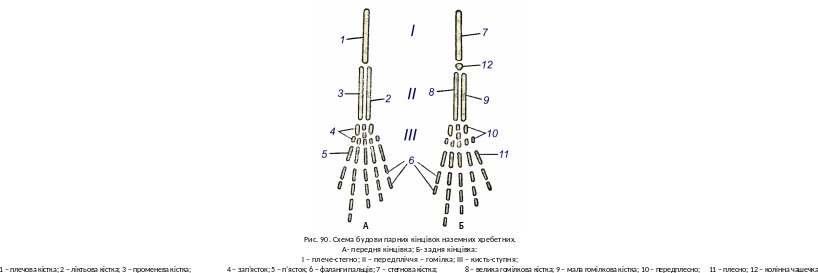 овна
редукція зябрової пόкришки призвела
до втрати під’язиковою дугою її основних
функцій.
овна
редукція зябрової пόкришки призвела
до втрати під’язиковою дугою її основних
функцій.
Парні кінцівки в земноводних за морфологією та функцією чітко відрізняються від парних плавців риб. Вони мають лише внутрішній скелет і побудовані за типом систем важелів, з’єднаних шарнірними суглобами. Ускладнення будови поясів забезпечує кінцівкам міцну опору, укріплюючи їх зв’язок з тулубом. Обидві пари кінцівок побудовані за єдиною схемою, спільною для всіх наземних хребетних (рис. 90).
Передня кінцівка
І. Плече; утворене плечовою кісткою (humerus).
II. Передпліччя (antebrachium); із двох кісток; променевої (radius) ліктьової (ulna).
III. Кисть (manus), розділяється на:
а) п’ясток (metacarpus), яка складається із п’яти видовжених кісток, розташованих в один ряд;
б) фаланги пальців (phalanges digitorum); звичайно розвивається 5 пальців; кожний з них має декілька фаланг.
Задня кінцівка
І. Стегно; утворене стегновою кісткою (femur).
II. Гомілка (arus); складається із двох кісток: великої гомілкової (tibia) і малої гомілкової (fibula).
III. Ступня (pes) підрозділяється:
а) передплесну (tarsus), яка складається із 9―10 кісток, розташованих в три ряди (3; 1―2; 5);
б) плюсну (metatarsus) із п’яти видовжених кісточок, розташованих в один ряд;
в) фаланги пальців (falanges digitorum); звичайно розвивається 5 пальців, кожний утворюється декількома фалангами.
П ояс
передніх кінцівок (плечовий) в земноводних
має вигляд напівкільця, що лежить у
товщі тулубової мускулатури. У безхвостих
скостеніла лопатка (scapula)
і коракоїд (coracoideum)
утворюють суглобну ямку для зчленування
з головкою плеча (рис. 91). До лопатки
причленовується широкий надлопатковий
хрящ (cartilago
suprascapularis)
і служить місцем прикріплення м’язів
спини. Перед коракоїдом лежить тонкий
паличкоподібний хрящ – прокоракоїд.
ояс
передніх кінцівок (плечовий) в земноводних
має вигляд напівкільця, що лежить у
товщі тулубової мускулатури. У безхвостих
скостеніла лопатка (scapula)
і коракоїд (coracoideum)
утворюють суглобну ямку для зчленування
з головкою плеча (рис. 91). До лопатки
причленовується широкий надлопатковий
хрящ (cartilago
suprascapularis)
і служить місцем прикріплення м’язів
спини. Перед коракоїдом лежить тонкий
паличкоподібний хрящ – прокоракоїд.
Знизу його прикриває тоненька покривна кістка – ключиця (clavicula). Ззаду місця злиття коракоїдів розташована кісткова грудина (sternum), яка має на кінці хрящове розширення. Перед ключицями лежить невеликий передгрудинник (episternum); передня його частина хрящова, а задня костеніє. Грудна клітка у земноводних відсутня. Хвостаті земноводні мають короткі ребра, які не доходять до грудини.
Т азовий
пояс складається із трьох елементів
(рис. 92), які з’єднуються один з одним,
утворюючи вертлужну ямку – місце
причленування головки стегна; у
безхвостих пояс майже повністю костеніє.
Довгі клубові кістки (ilium)
причленовуються до поперечних відростків
крижового хребця; сідничні кістки
(ischium)
зростаються одна з одною, знизу лежить
лобковий хрящ (cartilago
pubis).
У хвостатих земноводних сідничні і
клубові кістки – невеликих розмірів.
азовий
пояс складається із трьох елементів
(рис. 92), які з’єднуються один з одним,
утворюючи вертлужну ямку – місце
причленування головки стегна; у
безхвостих пояс майже повністю костеніє.
Довгі клубові кістки (ilium)
причленовуються до поперечних відростків
крижового хребця; сідничні кістки
(ischium)
зростаються одна з одною, знизу лежить
лобковий хрящ (cartilago
pubis).
У хвостатих земноводних сідничні і
клубові кістки – невеликих розмірів.
М’язова
система земноводних
істотно
відрізняється від такої у риб. Частина
тулубової мускулатури зберігає метамерну
будову. Однак спостерігається виразне
диференціювання: відособленні ділянки
м’язових сегментів зливаються в
стрічкоподібні м’язи. Різко зростає
маса м’язів кінцівок, яка представлена
складною їх системою – антагоністів:
згинаючих і розгинаючих суглоби.
Ускладнюється і спеціалізується
мускулатура ротової порожнини
 (жувальна, язика, дна ротової порожнини),
яка не лише бере участь в захоплені і
проковтуванні їжі, але й забезпечує
вентиляцію ротової порожнини і легень.
(жувальна, язика, дна ротової порожнини),
яка не лише бере участь в захоплені і
проковтуванні їжі, але й забезпечує
вентиляцію ротової порожнини і легень.
Т равна
система і живлення.
Всі сучасні земноводні в дорослому
стані твариноїдні. Вони поїдають різних
дрібних безхребетних: комах та їх
личинок, багатоніжок, павуків,
ракоподібних, червів і т.д. Водні види
можуть ловити молодь риби, а крупні із
них - земноводних та їх личинок, пташенят
водоплавних птахів, а також дрібних
гризунів, що попали у воду. Безногі
червуги ведуть риючий спосіб життя,
живляться ґрунтовими безхребетними,
окремі види селяться в термітниках та
мурашниках, поїдають їх жителів. Способи
добування їжі відносно одноманітні.
Безногі земноводні повільно пересуваючись
розшукують здобич з допомогою нюху і
дотику, хвостаті – з допомогою зору і
нюху. Здобич схоплюють щелепами; деякі
види хвостатих земноводних здобич
«приклеюють» до язика (наприклад,
печерна саламандра) (рис. 93). Безхвості
земноводні під час здобування їжі
орієнтуються переважно завдяки зору
і зрідка – нюху. Розшукують здобич,
пересуваючись повільними стрибками,
або частіше чатуванням. Захоплюють її,
викидаючи липкий язик, або захоплюють
щелепами; деякі види – шпорцева жаба,
зелені ропухи – інколи засовують здобич
в рот пальцями передніх лапок. Жаби,
які добре скачуть (види роду Rana
і ін.) здатні ловити також літаючих
комах. Малорухомі ропухи харчуються
переважно повзаючими комахами (жуки,
мурашки, гусінь та ін.).
равна
система і живлення.
Всі сучасні земноводні в дорослому
стані твариноїдні. Вони поїдають різних
дрібних безхребетних: комах та їх
личинок, багатоніжок, павуків,
ракоподібних, червів і т.д. Водні види
можуть ловити молодь риби, а крупні із
них - земноводних та їх личинок, пташенят
водоплавних птахів, а також дрібних
гризунів, що попали у воду. Безногі
червуги ведуть риючий спосіб життя,
живляться ґрунтовими безхребетними,
окремі види селяться в термітниках та
мурашниках, поїдають їх жителів. Способи
добування їжі відносно одноманітні.
Безногі земноводні повільно пересуваючись
розшукують здобич з допомогою нюху і
дотику, хвостаті – з допомогою зору і
нюху. Здобич схоплюють щелепами; деякі
види хвостатих земноводних здобич
«приклеюють» до язика (наприклад,
печерна саламандра) (рис. 93). Безхвості
земноводні під час здобування їжі
орієнтуються переважно завдяки зору
і зрідка – нюху. Розшукують здобич,
пересуваючись повільними стрибками,
або частіше чатуванням. Захоплюють її,
викидаючи липкий язик, або захоплюють
щелепами; деякі види – шпорцева жаба,
зелені ропухи – інколи засовують здобич
в рот пальцями передніх лапок. Жаби,
які добре скачуть (види роду Rana
і ін.) здатні ловити також літаючих
комах. Малорухомі ропухи харчуються
переважно повзаючими комахами (жуки,
мурашки, гусінь та ін.).
В земноводних ротовий отвір широкий веде в ротоглоткову порожнину, що переходить до стравоходу. В передній частині піднебіння у ротоглотковій порожнині відкриваються парні хоани – внутрішні отвори ніздрів, а неподалік щелепного суглоба знаходяться отвори євстахієвих труб, що ведуть в порожнину середнього вуха. У самців деяких жаб в кутах рота розміщені резонатори, або голосові мішки, які підсилюють звук. Дно задньої частини ротоглоткової порожнини підтримується хрящами гортанної щілини. В слизовій оболонці даху ротоглоткової порожнини розташовані слинні залози, які виділяють слизистий секрет, що не містить травних ферментів: він змочує порожнину і полегшує проковтування здобичі. Змочена слиною їжа переміщується в стравохід скороченням м’язів дна ротоглоткової порожнини, проковтуванню допомагають очі, які всовуються в ротоглоткову порожнину.
Стравохід (oesophagus) короткий, сильно розтягується, впадає в шлунок (gaster) (рис. 94, 6), він має товсті м’язові стінки. Від шлунка відходить дванадцятипала кишка (duodenum), яка непомітно переходить в тонку кишку (ipleum), що впадає в пряму кишку (rectum). Остання відкривається клоакою (cloaca). Трилопатева печінка (hepar) має в центральній лопаті жовчний міхур (рис. 94, 5); жовчна протока впадає в дванадцятипалу кишку. Між шлунком і дванадцятипалою кишкою лежить підшлункова залоза. Біля нижнього кінця шлунка розміщується селезінка (lien) – орган кровотворення та депо крові.
У шлунку виділяється фермент – пепсин, а у підшлунковій залозі – трипсин, амілаза й ліпаза.
Органи дихання та газообмін. Характерною особливістю земноводних є наявність багатьох органів дихання. В поглинанні кисню і виділенні вуглекислоти беруть участь: в личинок – шкіра, зовнішні та внутрішні зябра; у дорослих - легені, шкіра та слизова оболонка ротоглоткової порожнини. У деяких видів дорослих хвостатих земноводних зберігаються зовнішні зябра та недорозвинені або редуковані легені.
Дорослі земноводні дихають парними легенями, які мають вигляд порожнистих мішків, внутрішні стінки яких комірчасті (у деяких хвостатих вони відсутні).
Вентиляція легенів здійснюється за рахунок рухів дна ротоглоткової порожнини. При опусканні її повітря через відкриті ніздрі засмоктується в ротоглоткову порожнину, пізніше зовнішні отвори ніздрів закриваються, відкривається гортанна щілина і повітря із легень під тиском м’язів стінок тіла та внутрішніх органів витискається до ротоглоткової порожнини, де змішується з атмосферним повітрям. Дно ротоглоткової порожнини поступово піднімається, притискається до піднебіння, а змішане повітря нагнітається в легені. Потім гортанна щілина закривається і залишки змішаного повітря через ніздрі виштовхуються на зовні. В результаті обновлюється повітря в ротоглотковій порожнині і кров в капілярах її слизової насичується киснем.
Кровоносна система і кровообіг. Личинки земноводних мають одне коло кровообігу: в серці є одне передсердя та один шлуночок; є артеріальний конус, який розгалужується на чотири приносних зябрових артерій.
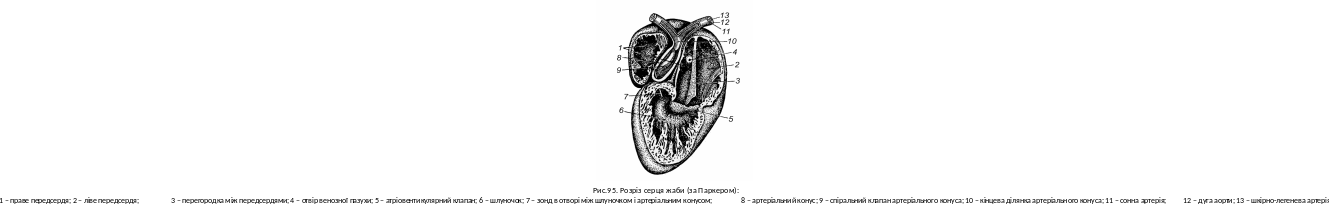
Серце дорослих земноводних трикамерне: два передсердя та один шлуночок (рис. 95). До правого передсердя примикає товстостінна венозна пазуха, від шлуночка відходить артеріальний конус. Таким чином, серце має п’ять відділів. Обидва передсердя відкриваються до шлуночка загальним отвором; тут розташовані атріовентрикулярні клапани (рис. 95, 5), які при скороченні шлуночка не пропускають кров назад в передсердя. Шлуночок має мускулисті вирости стінок, утворюючи ряд, з’єднаних одна з одною камер, що перешкоджає змішуванню крові. Артеріальний конус відходить від правої сторони шлуночка; всередині розташований довгий спіральний клапан (рис. 95, 9).
В
Рис. 96. Схема артеріальної системи жаби (більш артеріальна кров показана рідкою штриховкою, змішана густою штриховкою, венозна – чорним кольором):
1 – праве передсердя; 2 – ліве передсердя; 3 – шлуночок; 4 – артеріальний конус; 5 – шкірно-легенева артерія; 6 – легенева артерія; 7 – шкірна артерія; 8 – права дуга аорти; 9 – ліва дуга аорти; 10 – потилично-хребетна артерія; 11 – підключична артерія; 12 – спинна аорта; 13 – кишково-брижова артерія; 14 – сечостатеві артерії; 15 – загальна клубова артерія; 16 – загальна сонна артерія; 17 – внутрішня сонна артерія; 18 – зовнішня сонна артерія; 19 – легеня; 20 – печінка; 21 – шлуночок; 22 – кишечник; 23 – сім’янник; 24 –нирка.
 ід
артеріального конуса самостійними
отворами починаються три пари артеріальних
дуг. Першими від артеріального конуса
відходять права і ліва шкірно-легенева
артерія (arcus
pulmocutanea)
(рис. 96; 5) – гомологи IV
пари зябрових дуг личинок; вони
розпадаються на легеневу і шкірну
артерії. Другими – відходять системні
дуги (корені) аорти (arcus aortae) (рис. 96; 8, 9)
– гомологи ІІ пари зябрових дуг личинок.
Відділивши потилично-хребетну і
підключичну артерії, що забезпечують
кров’ю мускулатуру тулуба та передніх
кінцівок, вони зливаються під хребетним
стовбуром в спинну аорту (aorta
dorsalis)
(рис. 96; 12). Спинна аорта відділяє потужну
кишково-брижову артерію (забезпечує
кров’ю травну трубку); по інших
розгалуженнях спинної аорти кров йде
до решти органів та в задні кінцівки.
Останніми від артеріального конуса
відходять загальні сонні артерії
(acarotis
communis)
(рис. 96; 16) – гомологи І пари зябрових
дуг. Кожна із них розпадається на
зовнішні і внутрішні артерії (a.
c.
externa
et
interna).
ід
артеріального конуса самостійними
отворами починаються три пари артеріальних
дуг. Першими від артеріального конуса
відходять права і ліва шкірно-легенева
артерія (arcus
pulmocutanea)
(рис. 96; 5) – гомологи IV
пари зябрових дуг личинок; вони
розпадаються на легеневу і шкірну
артерії. Другими – відходять системні
дуги (корені) аорти (arcus aortae) (рис. 96; 8, 9)
– гомологи ІІ пари зябрових дуг личинок.
Відділивши потилично-хребетну і
підключичну артерії, що забезпечують
кров’ю мускулатуру тулуба та передніх
кінцівок, вони зливаються під хребетним
стовбуром в спинну аорту (aorta
dorsalis)
(рис. 96; 12). Спинна аорта відділяє потужну
кишково-брижову артерію (забезпечує
кров’ю травну трубку); по інших
розгалуженнях спинної аорти кров йде
до решти органів та в задні кінцівки.
Останніми від артеріального конуса
відходять загальні сонні артерії
(acarotis
communis)
(рис. 96; 16) – гомологи І пари зябрових
дуг. Кожна із них розпадається на
зовнішні і внутрішні артерії (a.
c.
externa
et
interna).
Венозна кров із заднього відділу тіла і задніх кінцівок збирається стегновими (v. femoralis) та сідничними (v .ischiadica) венами, останні зливаються в парні клубові або ворітні вени нирок (v. portae renalis) (рис. 97; 7), які розпадаються в нирках на капіляри, тобто утворюють ворітню систему нирок. Від правої і лівої стегнових вен відходять вени, що зливаються в непарну черевну вену (v. abdomilalis) (рис. 97; 8), яка проходить по черевній стінці до печінки, тут вона розпадається на капіляри. Венозна кров від всіх відділів кишечнику і шлунка збирається у велику ворітню вену печінки (v. portae hepatis), яка розпадаючись в печінці на капіляри (у всіх земноводних ворітня система печінки утворена черевною і ворітними венами). Капіляри нирок зливаються в численні виносні вени, які впадають в непарну задню порожнисту вену ( v. cava posterior); до неї впадають вени від статевих залоз. Задня порожниста вена проходить через печінку (кров із неї до печінки не проходить), і впадає у венозну пазуху.
А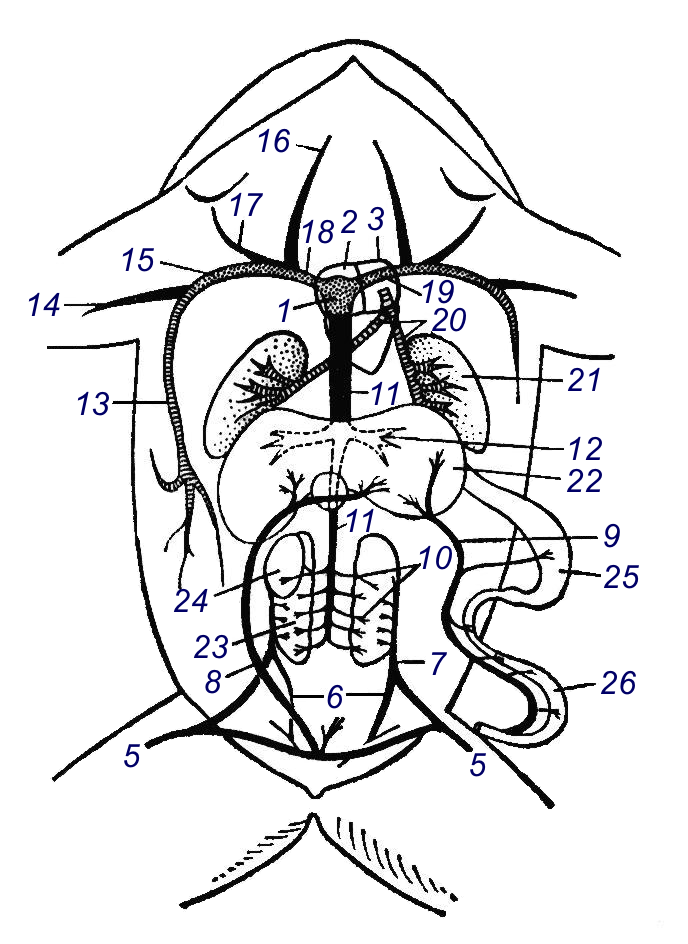
Рис. 97. Схема венозної системи жаби (більш артеріальна кров показана рідкою штриховкою, змішана – крапками, венозна – чорним кольором):
1 – венозна пазуха, 2 – праве передсердя, 3 – ліве передсердя, 4 – шлуночок, 5 – стегнова вена, 6 – сіднична вена, 7 – ворітна вена нирок, 8 – черевна вена, 9 – ворітна вена печінки, 10 – виносні ниркові вени, 11 – задня порожниста вена, 12 – печінкова вена, 13 – велика шкірна вена, 14 – плечова вена, 15 – підключична вена, 16 – зовнішня яремна вена, 17 – внутрішня яремна вена, 18 – права передня порожниста вена, 19 – ліва передня порожниста вена, 20 – легеневі вени, 21 – легеня, 22 – печінка, 23 – нирка, 24 – сім’яник, 25 – шлунок, 26 – кишечник.
ртеріальна кров, яка окислилась в шкірі, збирається у велику шкірну вену (v. cutanea magna) (рис. 97; 13), котра разом з плечовою веною, яка несе венозну кров із передньої кінцівки впадає в підключичну вену (v. subclavia). Підключичні вени зливаються з зовнішніми і внутрішніми яремними венами (v. jugularis externa et interna) в праву і ліву передні порожнисті вени (v. cava anterior dextra et sinistra) і впадають у венозну пазуху. Венозна кров по порожнистих венах із всіх відділів тіла і артеріальна кров, проходить по шкірних венах. Із венозної пазухи кров поступає в праве передсердя. Артеріальна кров із легень збирається в легеневі вени (v. pulmonalis) (рис. 97; 20) і впадає в ліве передсердя. Під час легеневого дихання в правому передсерді збирається змішана кров. Ліве передсердя заповнюється артеріальною кров’ю із легень. При одночасному скороченні передсердь кров поступає в шлуночок, де її перемішуванні заважають вирости його стінок: в правій частині шлуночка кров більш венозна, а у лівій – артеріальна. Артеріальний конус відходить від правої частини шлуночка. Тому при скороченні шлуночка в артеріальний конус спочатку поступає більш венозна кров, яка заповнює шкірно-легеневі артерії. При тривалому скорочені шлуночка тиск в артеріальному конусі зростає, спіральний клапан зрушується, відкриваються отвори дуг аорти, в які поступає змішана кров із центральної частини шлуночка. Коли шлуночок повністю скоротився, в конус поступає найбільш артеріальна кров із лівої половини шлуночка. Вона не може пройти в легенево-шкірні артерії і дуги аорти, тому що вони заповнені кров’ю. Напір крові, максимально стискуючи спіральний клапан, відкриває устя сонних артерій, куди потече, прямуючи в голову, артеріальна кров. Зменшення притоку кисню до головного мозку, може супроводжуватися пониженням загального рівня обміну речовин та впаданням тварини в оціпеніння.Органи виділення і водно-сольовий обмін. Вихід на сушу земноводних істотно проявився на характері водно-сольового обміну та на виведені із організму продуктів азотистого обміну. Личинки земноводних мають пронефричні нирки (переднирки). Під час метаморфозу розвиваються парні мезонефричні (тулубові) нирки. Нирки мають вигляд ущільнених тіл, що лежать з боків хребетного стовпа в ділянці поперекового хребця (рис. 98; 15). Від кожної нирки відходить сечопровід (вольфів канал), який впадає в клоаку. На дні клоаки є отвір, який веде в сильно розтягнутий сечовий міхур (vesica urinartia). На черевній поверхні нирок розташовані наднирники – залози внутрішньої секреції (glandula suprarenalis). Нирки отримують завдяки нирковим артеріям артеріальну кров із спинної аорти; значна кількість венозної крові проходить у нирки через ворітні вени нирок. Продукти білкового розпаду у дорослих земноводних виводяться переважно у вигляді сечовини (у личинок – у вигляді аміаку). При наповнені сечовиною сечового міхура скорочуються м’язи його стінок і концентрована сеча виводиться в клоаку і викидається назовні. Частина продуктів розпаду виділяється через шкіру. Велика роль шкіри у водному обміні. При високій вологості (під час руху через покриту росою траву) шкіра земноводних всмоктує воду, яка накопичилась в підшкірних лімфатичних порожнинах. За рахунок цього протягом короткого часу вага тварини зростає на 20―50%.
У видів, які пристосувались до життя у
більш сухих біотопах (ропухи), завдяки
ущільненню і частковому зроговінню
шкіри значно менше в тварин випаровується
води. У трав’яної жаби (Rana
temporaria),
що заселяє зволожені біотопи, шкіра
втрачає з кожного см2
поверхні до 30 мм3
води, а в пустелі зелена ропуха – Bufo
viridis
– лише 10 мм3.
Протеї, які постійно ведуть водний
спосіб життя гинуть при втраті 30―35%
води, жителі вологих біотопів (різні
жаби) – при втраті 40―50%
води, а живучі на деревах деякі квакші
– лише при втраті 70%. Водний обмін
речовин регулюється гормонами гіпофізу
та кортикоїдними гормонами – наднирників,
завдяки яким змінюється проникнення
через шкіру і стінки сечового міхура
води, інтенсивність фільтрації в нирках,
реабсорбція її та іонів солей в ниркових
канальцях.
видів, які пристосувались до життя у
більш сухих біотопах (ропухи), завдяки
ущільненню і частковому зроговінню
шкіри значно менше в тварин випаровується
води. У трав’яної жаби (Rana
temporaria),
що заселяє зволожені біотопи, шкіра
втрачає з кожного см2
поверхні до 30 мм3
води, а в пустелі зелена ропуха – Bufo
viridis
– лише 10 мм3.
Протеї, які постійно ведуть водний
спосіб життя гинуть при втраті 30―35%
води, жителі вологих біотопів (різні
жаби) – при втраті 40―50%
води, а живучі на деревах деякі квакші
– лише при втраті 70%. Водний обмін
речовин регулюється гормонами гіпофізу
та кортикоїдними гормонами – наднирників,
завдяки яким змінюється проникнення
через шкіру і стінки сечового міхура
води, інтенсивність фільтрації в нирках,
реабсорбція її та іонів солей в ниркових
канальцях.
Статева система, особливості розмноження. Земноводні роздільностатеві, мають парні статеві залози. Яєчники – ovarium є зернистої структури, підвішені на брижі і до весни заповнюють майже всю порожнину тіла. Біля яєчників розташовані багатолопатеві жирові тіла, в яких нагромаджуються запасні речовини для формування статевих продуктів під час зимової сплячки. Тонкі видовжені яйцепроводи – oviducti, представляють собою мюллерові канали. Кожний яйцепровід лійкою, розташованою в ділянці серця, відкривається в порожнину тіла; нижня частина лійки розширена і відкривається до клоаки. Функцію сечопровода виконують вольфові канали. Зрілі яйця через розрив стінок яєчника випадають в порожнину тіла, підхоплюються краями лійки, далі рухаються по яйцепроводах і покриваються слизовими білковими оболонками – виділеннями залоз стінок яйцепроводу, і нагромаджуються у відділах матки. Сім’яники круглі (testes) (рис. 98; 7), вони разом із жировими тілами звисають на брижі біля передніх країв нирок. Із кожного сім’яника виходить декілька тонких сім’явиносних канальців, що проходять в нирку і тут відкриваються у вольфів канал. У самців земноводних вольфів канал одночасно виконує функції сечопровода і сім’япровода. В нижній частині вольфового каналу утворюється здуття – сім’яний міхур (vesicula seminalis) який різко збільшується перед розмноженням і слугує резервуаром для сперматозоїдів. Вольфові канали отворами відкриваються в клоаку. У самців хвостатих земноводних зберігаються у вигляді тоненьких трубочок рудименти мюллерових каналів.
У безхвостих земноводних запліднення зовнішнє. З допомогою мозолів на кисті передніх кінцівок самець притримує самку. Самки відкладають ікру, а самці відразу скроплюють сім’яною рідиною. В частини хвостатих земноводних запліднення також зовнішнє, але своєрідне. Так, у родини кутозубів самка відкладає ікру в слизовий мішок, на котрий самець кладе сперматофор – пакет сперматозоїдів, облямованих слизовою оболонкою. Для більшості хвостатих земноводних запліднення внутрішнє. Самець тритонів відкладає сперматофор, а самка захоплює його краями клоаки, тут оболонка розчиняється і сперматозоїди, проникають в нижні ділянки яйцепроводу, де й запліднюють яйцеклітини. У безногих земноводних запліднення також внутрішнє: самець притискає вивернуту клоаку до зовнішнього отвору клоаки самки, вводячи в неї сім’яну рідину.
Більшість земноводних відкладають ікра у воду; в якій розвивається зародок. Личинка, яка вилупилась до метаморфозу веде водний спосіб життя. У жаб, квакш і інших безхвостих земноводних кожна ікринка обгорнута сильно набухаючою у воді слизовою оболонкою. Відкладені ікринки заклеюються в пакет, який плаває на воді. У ропух кладка яєць має вигляд тонких слизових шнурів, а джерлянки, тритони, саламандри відкладають поодинокі яйця, які приклеюються до підводних предметів.
У ропухи-повитухи – Alytes самець намотує джгути ікри на задні кінцівки і носить їх до вилуплення пуголовків. Квакші філомедузи – Phyllomedusa та деякі інші види тропічних деревних жаб відкладають ікру в листя, притримуючи задніми лапками, пізніше збивають кладку в пінистий пакет. Сурінамська піпа – P. pipa відкладає яйця в шкірні комірки на спині (рис. 93;.б) в яких проходить повний розвиток.
Для не багатьох видів земноводних властиве яйцеживородіння.
Плодючість різних видів земноводних коливається в широких межах. Так, зелені ропухи відкладають до 5―10 тис. ікринок, трав’яна жаба 800―4000, сіра ропуха – 1200―7000, тритони – від 100 до 500―600. Самка ринодерми Дарвіна відкладає лише 20―30 яєць, а червуга, яка охороняє кладку – 5―15.
У безхвостих земноводних сформована личинка звільняється від оболонок і за допомогою присоски (рис. 99; 18) прикріпляється до водних рослин. У хвостатих земноводних личинка вилуплюється більш сформованою, має краще розвинений хвіст та зовнішні зябра.
Т ривалість
личинкового періоду різна у різних
видів, вона залежить від погодних умов
і географічного розташування регіону
(клімат). У трав’яної жаби – Rana
temporaria
пуголовки в районі м. Києва завершують
метаморфоз у віці 35―40 днів, під Москвою
– через 45―55 днів і під Архангельськом
– через 60―70 днів. При несприятливих
погодних умовах (холодне літо), а саме
у північних частинах ареалу та в горах
пуголовки зимують у водоймах, завершують
метаморфоз на наступне літо. Такі
випадки зареєстровані під Москвою для
часничниці (часникової жаби) і озерної
жаби.
ривалість
личинкового періоду різна у різних
видів, вона залежить від погодних умов
і географічного розташування регіону
(клімат). У трав’яної жаби – Rana
temporaria
пуголовки в районі м. Києва завершують
метаморфоз у віці 35―40 днів, під Москвою
– через 45―55 днів і під Архангельськом
– через 60―70 днів. При несприятливих
погодних умовах (холодне літо), а саме
у північних частинах ареалу та в горах
пуголовки зимують у водоймах, завершують
метаморфоз на наступне літо. Такі
випадки зареєстровані під Москвою для
часничниці (часникової жаби) і озерної
жаби.
У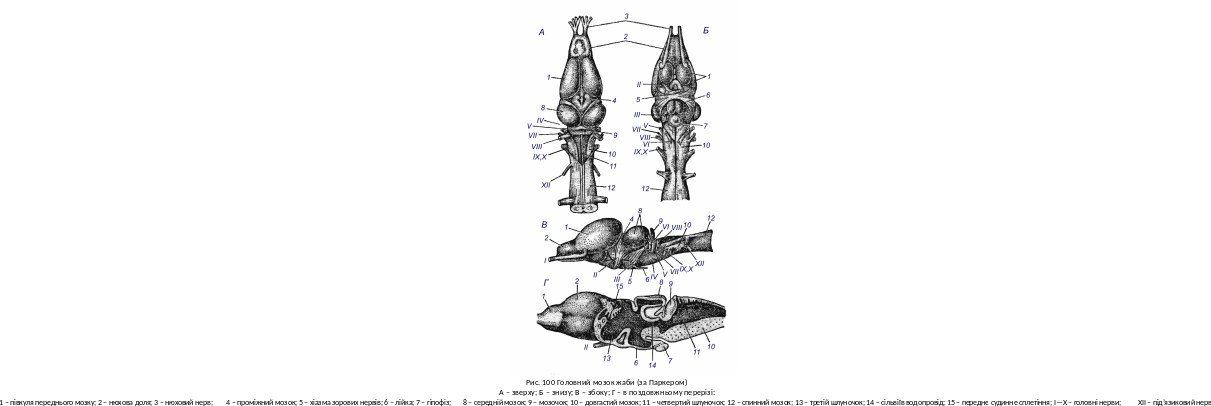 деяких хвостатих земноводних трапляється
неотенія – здатність личинок до
статевого розмноження. Вона добре
вивчена у живучої в Північній Америці
тигрової амбістоми – Ambystoma
tigrinum.
Личинки досить швидко завершують
метаморфоз у мілких, добре нагрітих
водоймах, з низьким вмістом кисню у
воді. Досягнувши довжини 5-9 см, вони
покидають водойми. В холодних водоймах
з високим вмістом кисню личинки добре
ростуть без метаморфозу; досягають
довжини 20-25 см, згодом у них розвиваються
статеві залози і вони приступають до
статевого розмноження. Згаданих личинок
розводять в лабораторних умовах під
назвою аксолотлів.
деяких хвостатих земноводних трапляється
неотенія – здатність личинок до
статевого розмноження. Вона добре
вивчена у живучої в Північній Америці
тигрової амбістоми – Ambystoma
tigrinum.
Личинки досить швидко завершують
метаморфоз у мілких, добре нагрітих
водоймах, з низьким вмістом кисню у
воді. Досягнувши довжини 5-9 см, вони
покидають водойми. В холодних водоймах
з високим вмістом кисню личинки добре
ростуть без метаморфозу; досягають
довжини 20-25 см, згодом у них розвиваються
статеві залози і вони приступають до
статевого розмноження. Згаданих личинок
розводять в лабораторних умовах під
назвою аксолотлів.
Можливо, деякі сиренові – Sirenidae, протеї – Proteidae і амфіуми – Amphiumidae це неотенічні личинки якихось саламандр, у котрих під час еволюції повністю зникала доросла фаза. Причиною такого могла бути висока смертність дорослих особин під впливом несприятливих погодних умов (похолодання, обледеніння і ін.), дією хижаків та інших факторів середовища. У безхвостих та безногих земноводних несприятливі погодні умови можуть викликати довгу затримку метаморфозу під час росту личинки.
Центральна нервова система і органи чуття. Перехід до наземного способу життя супроводжується перебудовою центральної нервової системи і органів чуття. Відносні розміри головного мозку земноводних у порівнянні з рибами помітно не зростають. Безхвості мають мозок трохи більший, ніж хвостаті. У сучасних земноводних помітно збільшуються відносні розміри переднього мозку, який розділений на дві півкулі (рис. 100) з самостійною порожниною – боковим шлуночком – у кожній з них.
Скупчення нервових клітин утворюють не лише смугасті тіла (corpora striata) на дні бокових шлуночків, а також тонкий шар в покрівлі півкуль – первинний мозковий купол – архіпаліум (archipallium). Нюхові долі слабо відмежовані від півкуль. Проміжний мозок зверху лише злегка прикритий сусідніми відділами. Зверху на ньому розташований епіфіз. Від дна проміжного мозку відходить лійка, до якої прилягає добре розвинутий гіпофіз. Середній мозок менший, ніж у кісткових риб. Мозочок малий і має вигляд невеликого валика, що лежить за середнім мозком на передньому краї ромбовидної ямки - порожнина четвертого шлуночка. Від головного мозку земноводних, як і у риб, відходять 10 пар головних нервів. Спинний мозок злегка сплющений, має плечове і поперекове потовщення, що пов’язане з відгалуженням могутніх нервових сплетінь, які іннервують передні і задні кінцівки. Симпатична нервова система у багатьох земноводних представлена двома стовбурами, що лежать з боків черевної сторони хребетного стовбура. Ганглії цих стовбурів з’єднані з спинномозковими нервами.
Органи чуття забезпечують орієнтування земноводних у воді та на суші. Для личинок і ведучих водний спосіб життя дорослих особин важливу роль відіграють органи бокової лінії, нюх, терморецепція, смак, слух та зір. Для видів земноводних з наземним способом життя основну роль в орієнтації відіграє зір.
Органи бокової лінії розкидані по - всьому тілі і лежать на поверхні шкіри. В останній розкидані дотикові тільця (скупчення чутливих клітин до яких підходять нерви).
Всі земноводні в епідермальному шарі шкіри мають вільні закінчення чутливих нервів. Вони сприймають температурні, больові і тактильні відчуття. В ротовій порожнині і на язиці містяться чутливі клітини оплетені нервовими закінченнями. Можливо вони не виконують функцію “смакових” рецепторів, а слугують органам дотику, дозволяють відчувати положення харчового об’єкту в ротовій порожнині. Про слабий смак у земноводних свідчить поїдання ними комах з різким запахом і їдким виділенням (мурашки, клопи, туруни та ін.).
Нюх у земноводних відіграє помітну роль. Для його забезпечення існують парні нюхові мішки. Зовнішні ніздрі відкриваються і закриваються за допомогою спеціальних м’язів. Внутрішніми ніздрями (хоанами) кожний мішок з’єднується з ротовою порожниною. Об’єм нюхових мішків і площа зайнята нюховим епітелієм, особливо велика у безногих (червуг) і в частини безхвостих (ропухи, деякі квакші) земноводних. Орган нюху функціонує лише у повітряному середовищі; у воді зовнішні ніздрі закриті. Значна роль нюху в орієнтації та пошуках їжі у риючих червуг. Хвостаті і безхвості земноводні розпізнають запах властивий конкретному біотопу, а також запах їжі. Відчуття нюху змінюється в різні сезони: дуже висока вона весною.
Органи зору добре розвинені у більшості наземних земноводних. Земноводні, що живуть в ґрунті (червуги) і підземних водоймах – (європейський протей, підземна саламандра – Typhlotriton spealaeus і декілька інших видів) – мають маленькі очі, які слабо просвічують через шкіру або їх не видно. Очі личинок не мають повік. Під час метаморфозу утворюються рухливі повіки – верхня і нижня – і миготлива перетинка, яка відокремлюється від нижньої повіки. В сітківці знаходяться палички і колбочки; у видів з присмерковою і нічною активністю переважають перші. У багатьох земноводних розвинуте сприйняття кольору.
Орган слуху у земноводних, в порівнянні з рибами істотно ускладнився. Перетинчастий лабіринт у земноводних, який міститься у капсулі внутрішнього вуха змінився мало. Виник новий відділ – середнє вухо, в якому розміщений апарат, що посилює сприйняття звукових коливань. Порожнина середнього вуха утворилась із рудименту зябрової щілини, розташованої між щелепами і під’язиковою дугами. Зовнішній отвір середнього вуха затягнутий тонкою пружною перетинкою. В порожнині знаходиться паличкоподібна кісточка – стремінце (stapes), яка одним кінцем впирається в барабанну перетинку, а другим – в овальне вікно, яке закрите меншою перегородкою.
Порівняльно-анатомічні і палеонтологічні дані показують, що стремінце утворилося із гіомандибуляра, або підвіска – верхнього елементу під’язикової дуги. Вузький канал – євстахієва труба (tuba Eustachii) з’єднує порожнину середнього вуха з ротовою порожниною, вирівнює тиск і запобігає розриви барабанної перетинки підчас сильних звуків.
Систематика класу разом із тими, що вимерли відносно проста.
Клас Земноводні, або Амфібії – Amphibia
Підклас Дугохребцеві – Apsidospondyli
+ Надряд Лабіринтодонти – Labyrinthodontia
+ Ряд Іхтіостегалії – Ichthyostegalia
+ Ряд Рахітомові –Rhachitomi
+ Ряд Стереоспондільні – Stereospondyli
+ Ряд Антракозаври - Anthracosauria
Надряд Стрибаючі – Salientia
+ Ряд Примітивні безхвості - Proanura
Ряд Безхвості – Anura (Ecaudata)
Підклас Тонкохребцеві – Lepospondyli
+ Ряд Нектрідіа – Nectridia
+ Ряд Аістопода - Aistopoda
+ Ряд Мікрозауріа - Microsauria
Ряд Хвостаті – Urodela (Caudata)
Ряд Безногі – Apoda
Вимерлі систематичні групи позначені знаком + (плюс).
Характеристика сучасних рядів земноводних
Земноводні є зараз наймалочисельнішим класом сучасних хребетних. В трьох його рядах нараховується близько 2500 видів