

8.6. Базальные ганглии
Базальные ганглии — совокупность расположенных в основании больших полушарий трех парных образований: бледного шара (pallidum), филогенетически более позднего образования — полосатого тела (striatum) и наиболее молодой части — ограды (claus-trum). Бледный шар состоит из наружного и внутреннего сегментов; полосатое тело пред-
137
ставлено хвостатым ядром (п. caudatus) и скорлупой (putamen). Ограда расположена между скорлупой и островковой корой. В функциональном отношении в базальные ганглии включают некоторые структуры промежуточного и среднего мозга: субталамичес-кое ядро и черное вещество (substantia nigra).
8.6.1. ФУНКЦИОНАЛЬНЫЕ СВЯЗИ БАЗАЛЬНЫХ ГАНГЛИЕВ
Афферентная импульсация поступает преимущественно в полосатое тело в основном из трех источников: от всех областей коры непосредственно и через таламус; от неспецифических интраламинарных ядер таламуса; от черного вещества. Среди эфферентных связей базальных ганглиев можно отметить три выхода:
-
от полосатого тела пути направляются к бледному шару; от бледного шара начина ется самый важный эфферентный тракт базальных ганглиев, идущий в таламус, в его релейные вентральные ядра, от них возбуждающий путь следует в двигатель ную кору (поля 4 и 6);
-
часть эфферентных волокон из бледного шара и полосатого тела идет к центрам ствола мозга (ретикулярная формация, красное ядро и далее — в спинной мозг), а также через нижнюю оливу в мозжечок;
-
от полосатоге тела тормозящие пути идут к черному веществу и после переключе ния — к ядрам таламуса.
Оценивая связи базальных ганглиев в целом, можно отметить, что эта структура является промежуточным звеном (станцией переключения), связывающей ассоциативную и, частично, сенсорную кору с двигательной корой.
В структуре связей базальных ганглиев выделяют несколько параллельно действующих функциональных петель, соединяющих базальные ганглии и кору больших полушарий [Александер Г. и др., 1986].
Скелетомоторная петля соединяет премо-торную, первичную моторную и соматосен-сорную области коры со скорлупой, импульсация из которой идет в бледный шар, черное вещество и далее через вентролатеральное ядро таламуса возвращается в премоторную область коры (поле 6). Считают, что эта петля служит для регуляции таких параметров движения, как амплитуда, сила, направление. При этом путь через черное вещество регулирует сокращение мышц лица.
Глазодвигательная петля соединяет области коры, контролирующие направление взгляда (поле 8 лобной коры и поле 7 теменной коры), с хвостатым ядром базальных ганглиев. Оттуда импульсация поступает в бледный шар и черное вещество, из которых она проецируется соответственно в ассоциативное медиодорсальное и релейное вентральное переднее ядра таламуса, а из них возвращается в лобное глазодвигательное поле 8. Эта петля участвует в регуляции, например, скачкообразных движений глаз (саккад). Предполагается существование также сложных петель, по которым импульсация из лобных ассоциативных зон коры поступает в структуры базальных ганглиев (хвостатое ядро, бледный шар, черное вещество) и через медиодорсальное и вентральное переднее ядра таламуса возвращается в ассоциативную лобную кору. Считают, что эти петли участвуют в регуляции высших психических функций мозга: контроля мотиваций, прогнозирования, познавательной (когнитивной) деятельности (см. Таламолобная система в разделе 8.7.3).
8.6.2. ФУНКЦИИ БАЗАЛЬНЫХ ГАНГЛИЕВ
Полосатое тело. Основными объектами функционального влияния полосатого тела являются бледный шар, черное вещество, таламус и моторная кора.
-
Влияние полосатого тела на бледный шар осуществляется преимущественно через тонкие тормозные волокна (медиатор ГАМК). Многие нейроны бледного шара дают смешанные ответы (ВПСП и ТПСП), а некоторые нейроны отвечают только ВПСП. Эти данные свидетельствуют о том, что поло сатое тело оказывает на бледный шар двоя кое действие — тормозящее и возбуждающее с преобладанием тормозящего.
-
Влияние на черное вещество. Между черным веществом и полосатым телом име ются двусторонние связи. Нейроны полоса того тела оказывают тормозящее влияние (медиатор ГАМК) на нейроны черного веще ства. В свою очередь нейроны черного веще ства модулируют влияние (медиатор—дофа мин) на кортико-стриарные каналы связи. До сих пор окончательно не установлен ха рактер этого влияния — тормозной, возбуж дающий или тот и другой. Дофаминергичес- ким путям в базальных ганглиях принадле жит важная роль в связи с двигательными расстройствами при паркинсонизме, при ко тором резко падает концентрация дофамина в обоих ядрах полосатого тела — хвостатом и
138
скорлупе. Кроме влияния на полосатое тело, черное вещество оказывает тормозящее действие на нейроны таламуса (медиатор ГАМК) и получает возбуждающие афферентные входы от субталамического ядра.
-
Влияние на таламус. Раздражение участ ков таламуса вызывает синхронизацию элек троэнцефалограммы: появление в ней высо коамплитудных ритмов, характерных для фа зы медленного сна. Эти ритмы можно вызвать раздражением не только таламуса, но и поло сатого тела. При этом из всех структур (черное вещество, миндалина, бледный шар), посыла ющих прямые пути к таламусу, только раздра жение полосатого тела вызывает появление в коре высокоамплитудных медленных ритмов. Напротив, разрушение полосатого тела нару шает эту реакцию и уменьшает время сна в цикле сон—бодрствование.
-
Влияние на моторную кору. Прямая сти муляция полосатого тела через хронически вживленные электроды вызывает относи тельно простые двигательные реакции: пово рот головы и туловища в сторону, противо положную раздражению, иногда сгибание ко нечности на противоположной стороне. Сти муляция некоторых зон полосатого тела вы зывает задержку текущей поведенческой дея тельности — двигательной, ориентировоч ной, пищедобывательной. Животное как бы «застывает» в одной позе. При этом на электроэнцефалограмме развиваются мед ленные высокоамплитудные ритмы. Раздра жение других точек полосатого тела приводит к подавлению ощущения боли.
Поражение полосатого тела (хвостатого ядра) дает достаточно характерную клиническую картину, в которой главными являются насильственные избыточные движения (гиперкинезы). Больной как бы не может справиться со своей мускулатурой. Эти гиперкинезы обозначаются как хорея и атетоз. При повреждении в эксперименте полосатого тела у млекопитающих стабильно развивается синдром гиперактивности: число бесцельных движений в пространстве увеличивается в 5— 7 раз. Опыты с разрушением полосатого тела показали его важную роль в организации поведения: животные с удаленной корой, но ин-тактными хвостатыми ядрами в упрощенном виде могут осуществлять некоторые формы поведения — передвижение в пространстве, поиск пищи, сохраняются ранее выработанные простые условные рефлексы. Дополнительное разрушение хвостатых ядер прекращает эти формы поведения.
Бледный шар. Получая из полосатого тела преимущественно тормозное и частично воз-
буждающее влияние, бледный шар оказывает модулирующее влияние на двигательную кору, мозжечок, ретикулярную формацию, красное ядро. При стимуляции бледного шара у животных преобладают элементарные двигательные реакции в виде сокращения мышц конечностей, шеи и лица. При этом выявлено влияние бледного шара на некоторые зоны гипоталамуса (центр голода и задний гипоталамус), отмечена активация пищевого поведения. Разрушение бледного шара сопровождается снижением двигательной активности: возникают адинамия, как бы отвращение ко всякому движению, сонливость, эмоциональная тупость, затрудняются осуществление имеющихся и выработка новых условных рефлексов.
Ограда. Электрическое раздражение различных зон ограды вызывает разнообразные соматические, вегетативные и поведенческие реакции, например пищевые, ориентировочные и эмоциональные, сокращение мышц туловища, головы, жевательные и глотательные движения и др. Двустороннее разрушение ограды вызывает слабо выраженные нарушения рефлексов позы, вегетативных реакций и условных рефлексов, например дифференци-ровки раздражителей.
В заключение отметим, что базальные ганглии — это прежде всего центры организации сложнейших видов моторной активности организма. Самые общие предположения о функциональной роли базальных ганглиев сводятся к следующему: допускается участие этого подкоркового комплекса в создании программ целенаправленных двигательных актов на основе информации, поступающей к ним практически из всех областей коры. Обработанная информация далее передается в ядра переднего таламуса, где она объединяется с информацией, поступающей из мозжечка. Совокупный комплекс возбуждения поступает в лобную кору, затем в двигательную кору, на нейронах которой конкретная программа реализуется, обеспечивая мозаичное возбуждение и торможение множества моторных нейронов, имеющих эфферентный выход на стволовые и спинальные двигательные центры. Базальные ганглии контролируют такие параметры движения, как сила, амплитуда, скорость и направление. Кроме участия в формировании двигательных актов, базальные ганглии включены в регуляцию цикла сон—бодрствование, механизмы формирования условных рефлексов, сложные формы восприятия внешней информации (например, осмысление текста).
139
8.7. НОВАЯ КОРА БОЛЬШОГО МОЗГА
Новая кора (неокортекс) представляет собой слой серого вещества общей площадью 1500—2200 см2, покрывающий большие полушария; она составляет около 72 % всей площади коры и около 40 % массы головного мозга. В коре имеется около 14 млрд нейронов, количество глиальных клеток примерно в 10 раз больше. Кора большого мозга является в филогенетическом плане наиболее молодой нервной структурой; у человека она осуществляет высшую регуляцию функций организма и психофизиологические процессы, обеспечивающие различные формы поведения.
8.7.1. СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ХАРАКТЕРИСТИКА
А. Нейронная организация новой коры. В направлении с поверхности в глубь коры различают 6 горизонтальных слоев.
I — молекулярный слой имеет очень мало клеток, но большое количество ветвящихся дендритов пирамидных клеток, формирую щих сплетение, расположенное параллельно поверхности. На этих дендритах образуют си напсы афферентные волокна, приходящие от ассоциативных и неспецифических ядер та- ламуса.
II — наружный зернистый слой составлен в основном звездчатыми клетками и частич но малыми пирамидными клетками. Волокна клеток второго слоя расположены преимуще ственно вдоль поверхности коры, образуя кортико-кортикальные связи.
-
— наружный пирамидный слой состоит в основном из пирамидных клеток средней величины. Аксоны этих клеток, как и зер нистые клетки II слоя, образуют кортико- кортикальные ассоциативные связи.
-
— внутренний зернистый слой по ха рактеру клеток (звездчатые клетки) и распо ложению их волокон аналогичен наружному зернистому слою. В этом слое имеют синап- тические окончания афферентные волокна, идущие от нейронов специфических ядер та- ламуса; здесь отмечена наибольшая плот ность капилляризации.
V — внутренний пирамидный слой обра зован средними и крупными пирамидными нейронами, причем в двигательной коре рас положены гигантские пирамидные клетки Беца. Аксоны этих клеток образуют эффе рентные кортико-спинальный и кортико- бульбарный двигательные тракты.
VI — слой полиморфных клеток образован преимущественно веретенообразными клетками, аксоны которых образуют кортико-та-ламические пути.
Оценивая в целом афферентные и эфферентные связи коры большого мозга, можно отметить, что в слоях I—IV происходит восприятие и обработка поступающих в кору сигналов. Напротив, покидающие кору эфферентные пути формируются преимущественно в V—VI слоях. Более детально деление коры на различные поля проведено на основе цитоархитектонических признаков (форма и расположение нейронов) К. Брод-маном (1909), который выделил 52 поля; многие из них характеризуются функциональными и нейрохимическими особенностями.
Гистологические данные показывают, что элементарные нейронные цепи, участвую щие в обработке информации, расположе ны перпендикулярно поверхности коры. Электрофизиологические исследования
[Маунткасл В., 1957] с погружением микроэлектродов перпендикулярно поверхности соматосенсорной коры показали, что все встречаемые на пути нейроны отвечают на раздражитель только одного качества (вида) (например, тактильный). Напротив, при погружении электрода под углом на его пути попадались нейроны разной модальности. Был сделан вывод, что в коре мозга имеются функциональные объединения нейронов, расположенные в цилиндрике диаметром 0,5—1,0 мм. Эти объединения были названы нейронными колонками. Они обнаружены в моторной коре, в различных зонах сенсорной коры. Соседние нейронные колонки могут взаимодействовать друг с другом.
Б. Кортиколизация функций — возрастание в филогенезе роли коры большого мозга в анализе и регуляции функций организма и подчинение коре нижележащих отделов ЦНС. Например, регуляция таких двигательных функций, как локомоция (прыжки, ходьба, бег) и выпрямительные рефлексы у низших позвоночных (амфибий), полностью обеспечивается стволом мозга, удаление больших полушарий практически их не изменяет. У кошек перерезка ствола между средним и промежуточным мозгом существенно не влияет на выпрямительные рефлексы, но нарушает в остром периоде локомоцию, которая в дальнейшем частично восстанавливается. Выключение коры больших полушарий у обезьян и у человека приводит к потере не только локомоции, но и выпрямительных рефлексов.
140
В. Локализация функций в коре большого мозга интенсивно изучается в клинической и экспериментальной медицине, начиная с середины XIX в. При разработке этой проблемы были сформулированы две противоположные по смыслу концепции: узкого лока-лизационизма и функциональной равноценности (эквипотенциальности) различных корковых структур. Современная концепция локализации функций базируется на принципе многофункциональности (но не равноценности) корковых полей. Свойство мульти-функциональности позволяет той или иной корковой структуре включаться в обеспечение различных форм деятельности, реализуя при этом основную, генетически присущую ей функцию [Адрианов О.С, 1976]. Степень мультифункциональности различных корковых структур не одинакова. В полях ассоциативной коры она выше, чем в первичных сенсорных полях. В основе мультифункциональности лежат многоканальность поступления в кору мозга афферентного возбуждения, перекрытия афферентных возбуждений, особенно на таламическом и корковом уровнях, модулирующее влияние различных структур (например, неспецифического тала-муса, базальных ганглиев) на корковые функции, взаимодействие корково-подкор-ковых и межкорковых путей проведения возбуждения. Результаты изучения локализации функций позволили ученым в середине XX в. разделить кору головного мозга на различные функциональные зоны (картирование коры). Детальные функциональные карты (например, К. Клейста, 1959) используются в неврологической практике, однако надо иметь в виду упрощенный характер этих схем. Одним из наиболее известных вариантов функционального разделения коры большого мозга является выделение в ней сенсорной, ассоциативной и двигательной областей.
8.7.2. СЕНСОРНЫЕ ОБЛАСТИ КОРЫ
Это зоны, в которые проецируются сенсорные раздражители (син.: проекционная кора, корковые отделы анализаторов). Они расположены преимущественно в теменной (поля 1-3), височной (поля 21, 22, 41, 42) и затылочной (поля 17—19) долях. Афферентные пути в сенсорную кору поступают преимущественно от релейных сенсорных ядер тала-муса — вентральных задних латерального (п. VPL) и медиального (п. VPM).
Зоны сенсорной коры, раздражение или разрушение которых вызывает четкие и по-
стоянные изменения чувствительности организма, называются первичными сенсорными областями (ядерные части анализаторов, по И.П. Павлову). Они состоят преимущественно из мономодальных нейронов и формируют ощущения одного качества. В первичных сенсорных зонах обычно имеется четкое пространственное (топографическое) представительство частей тела, их рецептор-ных полей. Вокруг первичных сенсорных зон находятся менее локализованные вторичные сенсорные зоны, полимодальные нейроны которых отвечают на действие нескольких раздражителей.
Важнейшими сенсорными областями являются теменная кора постцентральной извилины и соответствующая ей часть парацентральной дольки на медиальной поверхности полушарий (поля 1—3). Эту зону обозначают как соматосенсорную область I. Здесь имеется проекция кожной чувствительности противоположной стороны тела от тактильных, болевых, температурных рецепторов, интероцептивной чувствительности и чувствительности опорно-двигательного аппарата от мышечных, суставных, сухожильных рецепторов. Проекция головы и верхних отделов туловища находится в нижнелатеральных участках постцентральной извилины, проекция нижней половины туловища и ног — в верхнемедиальных зонах извилины, проекция нижней части голени и стоп расположена в коре парацентральной дольки на медиальной поверхности полушарий (рис. 8.7), при этом проекции наиболее чувствительных участков (язык, губы, гортань, пальцы) имеют относительно большие зоны по сравнению с другими частями тела. Предполагается, что в соматосенсорной области I в зоне тактильной чувствительности языка расположена проекция и вкусовой чувствительности.
Кроме соматосенсорной области I, выделяют меньших размеров соматосенсорную область II, расположенную на границе пересечения центральной борозды с верхним краем височной доли, в глубине латеральной борозды. Степень локализации частей тела здесь выражена хуже; проекция лица находится спереди и ниже, рук — центрально, ног — сзади и выше. Функции области II плохо изучены. Известно, что сигналы в эту область поступают с обеих сторон тела и от других сенсорных областей мозга, например зрительных и слуховых. Раздражение области II приводит к сложным движениям тела; предполагают ее роль в сенсорном контроле движения.
141
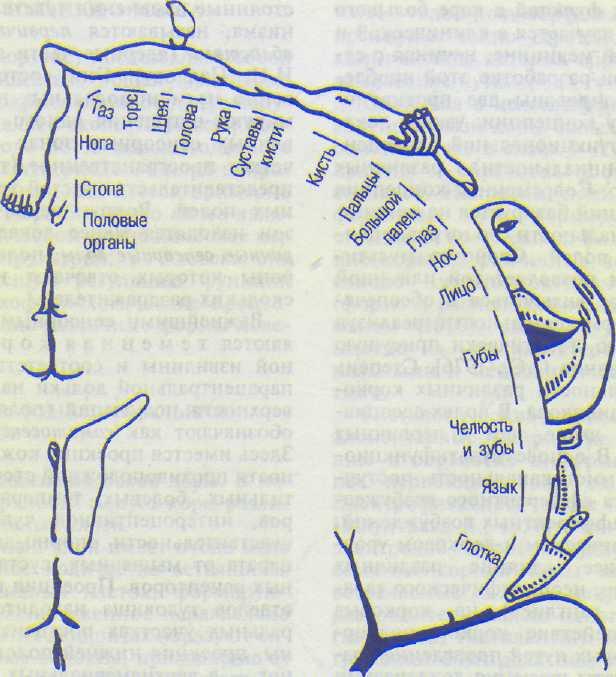
Хорошо изученной первичной проекционной зоной является слуховая кора (поля 41, 42), которая расположена в глубине латеральной борозды (кора поперечных височных извилин Гешля). В этой зоне в ответ на раздражение слуховых рецепторов кортиева органа формируются звуковые ощущения, различающиеся по громкости, тону и другим характеристикам. Здесь имеется четкая топическая проекция: в разных участках коры представлены различные участки кортиева органа. К проекционной коре височной доли относится также центр вестибулярного анализатора в верхней и средней височных извилинах (поля 20 и 21). Обработанная сенсорная информация используется для формирования «схемы тела» и регуляции функций мозжечка (височно-мостомозжечковый путь).
Важнейшая первичная проекционная область новой коры расположена в затылоч-
ной коре — это первичная зрительная область (кора части клиновидной извилины и язычковой дольки, поле 17). Здесь имеется топическое представительство рецепторов сетчатки. Каждой точке сетчатки соответствует свой участок зрительной коры, при этом зона желтого пятна имеет сравнительно большую зону представительства. В связи с неполным перекрестом зрительных путей в зрительную область каждого полушария проецируются одноименные половины сетчатки. Наличие в каждом полушарии проекции сетчатки обоих глаз является основой бинокулярного зрения. Раздражение коры поля 17 приводит к возникновению световых ощущений. Около поля 17 расположена кора вторичной зрительной области (поля 18 и 19). Нейроны этих зон полимодальны и отвечают не только на световые, но и на тактильные и слуховые раздражители.
142
В данной зрительной области происходит синтез различных видов чувствительности, возникают более сложные зрительные образы и их опознание. Раздражение этих полей вызывает зрительные галлюцинации, ауру (навязчивые ощущения), движение глаз.
Основная часть информации об окружающей среде и внутренней среде организма, поступившая в сенсорную кору, передается для дальнейшей ее обработки в ассоциативную кору.
8.7.3. АССОЦИАТИВНЫЕ ОБЛАСТИ КОРЫ
Ассоциативная (межсенсорная, межанализаторная кора) включает участки новой коры большого мозга, которые расположены рядом с сенсорными и двигательными зонами, но не выполняют непосредственно чувствительных или двигательных функций. Гра-
ницы этих областей обозначены недостаточно четко, неопределенность преимущественно связана со вторичными (высшими) проекционными зонами, функциональные свойства которых являются переходными между свойствами первичных проекционных и ассоциативных зон. Ассоциативная кора является филогенетически наиболее молодой частью новой коры, получившей наибольшее развитие у приматов и человека. У последнего она составляет около 50 % всей коры и 70 % неокортекса.
Основной физиологической особенностью нейронов ассоциативной коры, отличающей их от нейронов первичных зон, является по-лисенсорность (полимодальность): они отвечают, как правило, не на один, а на несколько раздражителей — зрительные, слуховые, кожные и др. Полисенсорность нейронов ассоциативной коры создается кортико-кортикальными связями с разными проекционными зо-
143
нами, связями с ассоциативными ядрами та-ламуса. В результате этого ассоциативная кора представляет собой своеобразный коллектор различных сенсорных возбуждений.
По таламокортикальным проекциям выделяют две ассоциативные системы мозга: тала-мотеменную и таламолобную. Делаются попытки выделить таламовисочную систему.
Таламотеменная система представлена ассоциативными зонами теменной коры (поля 5, 7, 40), получающими основные афферентные входы от задней группы ассоциативных ядер таламуса (латеральное заднее ядро и подушка). Теменная ассоциативная кора имеет эфферентные выходы на ядра таламуса и гипоталамуса, в моторную кору и ядра экстра-пирамидной системы. Основными функциями таламотеменной системы являются гно-зис, формирование «схемы тела» и праксис. Под гнозисом понимают функцию различных видов узнавания — формы, величины, значения предметов, понимание речи, познание процессов, закономерностей и др. К гностическим функциям относится оценка пространственных отношений (например, взаимного расположения предметов). В теменной коре выделяют центр стереогнозиса, расположенный сзади от средних отделов постцентральной извилины (поля 7, 40, частично 39), обеспечивающий способность узнавания предметов на ощупь. Вариантом гностической функции является формирование в сознании трехмерной модели тела («схемы тела»), центр «схемы тела» расположен в поле 7 теменной коры. Под праксисом понимают целенаправленное действие. Центр праксиса находится в надкраевой извилине (поля 39 и 40 доминантного полушария); он обеспечивает хранение и реализацию программы двигательных автоматизированных актов (например, рукопожатие, причесывание, зажигание спички и др.).
Таламолобная система представлена ассоциативными зонами лобной коры (поля 9— J4), имеющими основной афферентный вход от ассоциативного медиодорсального ядра таламуса, других подкорковых ядер.
Основная роль лобной ассоциативной коры сводится к инициации базовых системных механизмов формирования функциональных систем целенаправленных поведенческих актов (П.К. Анохин). Реализация этого сложнейшего процесса основана на участии нейронного субстрата лобной коры в решении следующих частных задач.
• Взаимоувязка возбуждений доминирующей мотивации с возбуждениями сенсор-
ного и биологического качества, поступающими в кору мозга из внешней среды.
-
Обеспечение прогнозирования в достиже нии значимых для жизнедеятельности по лезных результатов.
-
Самоконтроль эффективности поведен ческих актов путем сравнения достигну тых конечных результатов с афферентны ми моделями этих результатов в аппарате предвидения — акцепторе результатов действия (П.К. Анохин).
В результате операции префронтальной лоботомии, при которой лобные доли по медицинским показаниям изолируются, наблюдаются выраженные изменения поведения, в котором обычно доминируют эмоциональные расстройства, неадекватность поступков и действий, особенно в изменившихся условиях.
Концепция таламовисочной системы не получила еще достаточной научной проверки. Надо отметить, что некоторые ассоциативные центры (например, стереогнозиса, праксиса) включают в себя и участки височной коры (поле 39). В височной коре расположен слуховой центр речи Вернике, находящийся в задних отделах верхней височной извилины (поля 22, 37 и 42 левого доминантного полушария). Этот центр обеспечивает речевой гнозис — распознание и хранение устной речи, как собственной, так и чужой. В средней части верхней височной извилины (поле 22) находится центр распознания музыкальных звуков и их сочетаний. На границе височной, теменной и затылочной долей (поле 39) локализован центр чтения письменной речи, обеспечивающий распознание и хранение образов письменной речи.
Психические функции, осуществляемые ассоциативной корой, инициируют поведение организма, обязательным компонентом которого являются произвольные целенаправленные движения, осуществляемые при обязательном участии двигательной коры.
8.7.4. ДВИГАТЕЛЬНЫЕ ОБЛАСТИ КОРЫ
Представление о двигательной коре большого мозга начало формироваться с 80-х годов XIX в., когда было показано, что электрическое раздражение некоторых корковых зон у животных вызывает движение конечностей противоположной стороны [Фрич Г., Гит-циг Е., 1870]. В двигательной коре выделяют первичную и вторичную моторные области. В первичной двигательной коре
144
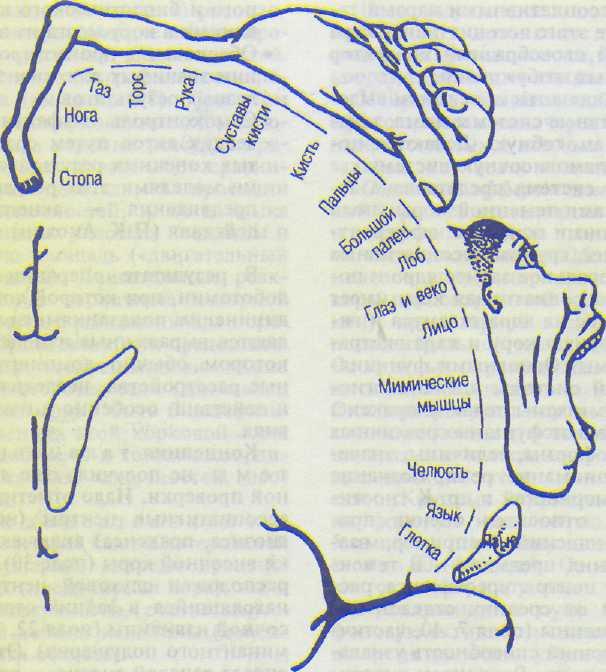
(прецентральная извилина, поле 4) расположены нейроны, иннервирующие мотонейроны мышц лица, туловища и конечностей. В ней имеется четкая топографическая проекция мышц тела (см. рис. 8.7). При этом проекции мышц нижних конечностей и туловища расположены в верхних участках пре-центральной извилины и занимают сравнительно небольшую площадь, а проекции мышц верхних конечностей, лица и языка расположены в нижних участках извилины и занимают большую площадь («двигательный человечек» Пенфилда). Двигательные реакции на раздражение первичной моторной коры осуществляются с минимальным порогом (высокая возбудимость): они представлены элементарными сокращениями мышц противоположной стороны тела (для мышц головы сокращение может быть билатеральным). При поражениях этой корковой зоны утрачивается способность к тонким координированным движениям конечностей, особенно пальцев рук.
Вторичная двигательная ко-р а (поле 6) расположена как на латеральной поверхности полушарий, впереди прецент-ральной извилины (премоторная кора), так и на медиальной поверхности, соответствующей коре верхней лобной извилины (дополнительная моторная область).
Вторичная двигательная кора в функциональном плане имеет главенствующее положение по отношению к первичной двигательной коре, осуществляя высшие двигательные функции, связанные с планированием и координацией произвольных движений. Здесь в наибольшей степени регистрируется медленно нарастающий отрицательный потенциал готовности, возникающий примерно за 1 с до начала движения. Кора поля 6 получает основную часть импульсации от базальных ганглиев и мозжечка, участвует в перекодировании информации о плане сложных движений. Раздражение коры поля 6 вызывает сложные координированные движения, например поворот головы, глаз и туловища в противоположную сторону, содружественные сокращения сгибателей или разгибателей на противоположной стороне. В премоторной коре расположены двигательные центры, связанные с социальными функциями человека: центр письменной речи в заднем отделе средней лобной извилины (поле 6), центр моторной речи Брока в заднем отделе нижней лобной извилины (поле 44), обеспечивающие речевой праксис, а также музыкальный моторный центр (поле 45), тональности речи.
При раздражении дополнительной моторной области возникают двигательные реакции формирования позы, ритмические и некоординированные движения.
Нейроны двигательной коры получают афферентные входы через таламус от мышечных, суставных и кожных рецепторов, от базальных ганглиев и мозжечка.
Основные эфферентные связи двигательной коры осуществляются через пирамидные и экстрапирамидные тракты, начинающиеся от гигантских пирамидных клеток Беца и менее крупных пирамидных клеток V слоя коры прецентральной извилины (60 % волокон), премоторной коры (20 % волокон) и постцентральной извилины (20 % волокон). От крупных пирамидных нейронов формируются преимущественно пирамидные пути. Они иннервируют крупные (фазические, высокопороговые) а-мотонейроны в моторных центрах ствола и спинного мозга, регулирующие движение тела и его частей в пространстве. Крупные пирамидные клетки имеют фоновую импульсную активность около 5 имп/с, которая при движениях увеличивается до 20—30 имп/с. От мелких пирамидных клеток формируются преимущественно экстрапирамидные пути. Эти клетки имеют фоновую активность около 15 имп/с, которая во время движения увеличивается или уменьшается; они иннервируют мелкие (тонические, низкопороговые) а-мотонейроны в стволовых и спинальных двигательных центрах и регулируют тонус мышц и позу.
Волокна пирамидного пути оканчиваются на а-мотонейронах двигательных ядер III— VII и IX—XII черепных нервов (кортико-бульбарный тракт) или спинальных двигательных центрах (кортико-спинальные тракты). Через двигательную кору и пирамидные пути осуществляются произвольные простые движения и сложные целенаправленные двигательные программы (например, профессиональные навыки), формирование которых начинается в базальных ганглиях и мозжечке и заканчивается во вторичной моторной коре. Большинство волокон пирамидных путей осуществляет перекрест, однако небольшая часть волокон остается неперекре-щенными, что способствует компенсации нарушения функции движения при односторонних поражениях. Через пирамидные тракты осуществляет свои функции и премоторная кора. К ним относятся двигательные навыки письма, сочетанные повороты головы, глаз и туловища, речевые функции (речедви-гательный центр Брока, поле 44). В регуляции письменной и особенно устной речи
145
имеется выраженная асимметрия больших полушарий мозга: у 95 % правшей и у 70 % левшей устная речь контролируется левым полушарием.
К корковым экстрапирамидным путям относятся кортико-рубральные и кортико-ре-тикулярные тракты, начинающиеся приблизительно от зон, которые дают начало пирамидным путям. Волокна кортико-рубрально-го тракта оканчиваются на нейронах красных ядер среднего мозга, от которых далее идут руброспинальные тракты. Волокна кортико-ретикулярных трактов оканчиваются на нейронах медиальных ядер ретикулярной формации моста (от них идут медиальные ретику-лоспинальные тракты) и на нейронах ретикулярных гигантоклеточных ядер продолговатого мозга, от которых начинаются латеральные ретикулоспинальные тракты. Через эти тракты осуществляется регуляция тонуса и позы, обеспечивающих точные, целенаправленные движения. Корковые экстрапирамидные тракты являются компонентом экстрапирамидной системы головного мозга, к которой относятся мозжечок, базальные ганглии, моторные центры ствола; она осуществляет регуляцию тонуса, позы, координацию и коррекцию движений. Поскольку кортико-пирамидные пути отдают многочисленные коллатерали к структурам экстрапирамидной системы, обе системы работают в функциональном единстве, и некоторые авторы [Шмидт Р., 1985] считают нецелесообразным их разделение. О функциональном единстве этих систем свидетельствуют и опыты по перерезке пирамидных путей у приматов. У таких животных не было отмечено грубых нарушений движений: сохранялись стояние, ходьба, двигательные условные рефлексы, сложные программированные движения. Не восстанавливались только «специфические» функции пирамидных путей — независимые, точные движения пальцев рук и контроль силы этих движений.
Определяющую роль в структурном обеспечении двигательных актов играют следующие образования:
-
фронтальная кора, выступающая в качест ве инициатора замысла действия;
-
ассоциативная теменная кора, выполняю щая функции интеграции гетеромодаль- ных возбуждений, а также командные функции в отношении ряда параметров движения;
-
базальные ганглии, мозжечок, возможно, выполняющие функции накопителей «двигательных программ»;
• моторная кора, моторные центры ствола, спинного мозга, обеспечивающие выбор мышечных эффекторов в ходе реализации программы двигательного акта и управления двигательным процессом на базе обратной афферентации (П.К. Анохин, А.С. Батуев).
8.7.5. МЕЖПОЛУШАРНЫЕ ВЗАИМООТНОШЕНИЯ
У человека межполушарные взаимотношения проявляются в двух главных формах — функциональной асимметрии больших полушарий и совместной их деятельности.
Функциональная асимметрия полушарий является важнейшим психофизиологическим свойством головного мозга человека. Ее исследование началось в середине прошлого века, когда французские медики М. Дакс (1836) и П. Брока (1861) показали, что нарушение речи человека возникает при поражении коры нижней лобной извилины, как правило, левого полушария. Несколько позже немецкий психиатр К. Вернике (1874) обнаружил в коре заднего отдела верхней височной извилины левого полушария слуховой центр речи, поражение которого приводит к нарушению понимания устной речи. Эти результаты и наличие моторной асимметрии (праворукость) сформировали концепцию, согласно которой для человека характерно левополушарное доминирование, образовавшееся эволюционно в результате трудовой деятельности и являющееся специфическим свойством его мозга. Однако в XX в. в результате применения различных методических подходов, особенно при исследовании больных с расщепленным мозгом (перерезка мозолистого тела), было показано, что по ряду психофизиологических функций доминирует не левое, а правое полушарие, и возникла концепция частичного доминирования полушарий [Сперри Р., 1981].
Выделяют психическую, сенсорную и моторную межполушарные функциональные асимметрии мозга. При исследовании речи было показано, что словесный информационный канал контролируется левым полушарием, а несловесный канал (интонация) — правым. Абстрактное мышление и сознание связаны преимущественно с левым полушарием. При выработке условного рефлекса в начальной фазе доминирует правое полушарие, а во время упрочения рефлекса — левое. Правое полушарие осуществляет обработку информации одновременно, синтетически, по
146
принципу дедукции, при этом лучше воспринимаются пространственные и относительные признаки предметов. Левое полушарие производит обработку информации последовательно, аналитически, по принципу индукции, лучше воспринимает абсолютные признаки предметов и временные отношения. В эмоциональной сфере правое полушарие обусловливает преимущественно более древние, отрицательные эмоции, контролирует проявления сильных эмоций, в целом оно более «эмоционально». Левое полушарие обусловливает в основном положительные эмоции, контролирует проявление более слабых эмоций.
В сенсорной сфере роль правого и левого полушарий лучше всего проявляется при зрительном восприятии. Правое полушарие воспринимает зрительный образ целостно, сразу во всех подробностях, легче решает задачу различения предметов и опознания бессмысленных предметов, которые трудно описать словами, создает предпосылки конкретно-чувственного мышления. Левое полушарие оценивает зрительный образ расчлененно, аналитически, при этом каждый признак (форма, величина и др.) анализируется отдельно, легче опознаются знакомые предметы и решаются задачи сходства предметов; зрительные образы лишены конкретных подробностей и имеют высокую степень абстракции; создаются предпосылки логического мышления.
Моторная асимметрия связана с тем, что мышцы конечностей и туловища одной стороны тела контролируются моторной корой противоположного полушария (мышцы лица контролируются обоими полушариями).
Функциональная асимметрия больших полушарий, обеспечивая новый, более высокий уровень регуляции сложных функций мозга, вместе с тем повышает требования к совмещению деятельности двух полушарий.
Парность в деятельности больших полушарий обеспечивается наличием комиссураль-ной системы — мозолистого тела, передней, задней, гиппокампальной и хабенулярной комиссур, межбугрового сращения, которые анатомически соединяют два полушария головного мозга. Результаты, свидетельствующие о переносе возбуждения из одного полушария в другое, получены преимущественно с использованием электрофизиологических методик и метода условных рефлексов.
Основные факты, полученные с помощью электрофизиологических методик, показали, что возбуждение из участка раздражения одного полушария передается через комиссу-
ральную систему не только в симметричный участок другого полушария, но также и в несимметричные участки коры (например, из зрительной коры одного полушария в моторную кору противоположного полушария). Морфологические исследования также показали, что, кроме поперечных волокон мозолистого тела, связывающих симметричные участки коры, есть продольные и вертикальные волокна, связывающие несимметричные корковые поля.
Использование метода условных рефлексов показало, что если рефлекс вырабатывается, например, на тактильное раздражение кожи одной стороны тела, то он легко воспроизводится при таком же раздражении симметричных участков кожи другой стороны тела, т.е. в процессе выработки рефлекса происходит «перенос» временной связи в другое полушарие. Предварительная перерезка мозолистого тела прекращала (или сильно затрудняла) «перенос» условного рефлекса. Вместе с тем с помощью метода условных рефлексов было показано, что перерезка всех комиссур больших полушарий полностью не исключает возможности межполушарного взаимодействия при выработке временной связи. Эти результаты свидетельствуют о том, что элементарные формы взаимодействия двух полушарий могут осуществляться через подкорковые и стволовые структуры.