

Раздел 1. Смерть младенца вследствие трудных родов
Больная Б. Наблюдалась во время беременности собственным доктором и в стационаре. Ее последний ребенок родился весом 4,4 кг, во время родов была выявлена небольшая дистоция плеча. Больная была осмотрена консультантом общества акушерок в сроке 38 недель, так как ребенок был слишком крупным для этого срока. На ультразвуковом исследовании вес ребенка был определен в 4,5 кг. Пальпация и ультразвуковое исследование проводились с учетом анамнеза предыдущих родов. Во-первых, беременность не будет продолжаться более 6 дней до начала родов (а не 12-14 дней, как обычно). Во-вторых, было записано, что в случае трудных родов не будут использоваться инструментальные методы. В-третьих, прогнозировалась дистоция плеча и была сделана соответствующая запись для предупреждения акушеров.
Хронология:
|
5:55 |
Больная Б. госпитализирована с явлениями разрыва оболочек. Вскоре после этого началась родовая деятельность. |
|
6:50 |
При вагинальном исследование выявлено расширение шейки на 3 см. Сердцебиение плода отслеживалось с помощью внешнего допплеровского датчика. На этом этапе больная попросила эпидуральную аналгезию, но анестезиолог на этот момент был занят в отделении интенсивной терапии. Роды у больной протекали быстро, поэтому эпидураль-ная блокада не была выполнена. |
|
7:15 |
В связи с тем, что акушерки потеряли возможность мониторировать сердцебиение плода из-за размеров и напряжения матки, на голову плода был помещен электрод. Полученная кривая показывала нормальное сердцебиение плода. |
|
7:50 |
Проведено вагинальное исследование. Шейка расширена до 6 см. Сердцебиение плода нормальное. Введен петидин. |
|
8:05 |
Шейка полностью раскрыта. Начало выталкивания. Мать не может действовать вместе с персоналом из-за боли. |
|
8:14 |
Электрод с головы плода удален во время прорезывания. Последние данные о сердцебиении плода показали значительное урежение. Родовая деятельность отсутствовала, голова плода оставалась неподвижной. Вновь присоединенный внешний допплеровский датчик показал частоту сердечных сокращений плода 160-170 ударов в минуту. |
|
8:33 |
Потребовалась медицинская помощь. Акушер и дежурный консультант немедленно пришли и быстро диагносцировали дистоцию плеча. Они провели маневр Макробертса, применили надлобковое надавливание и ребенок родился в 8:39. |
Состояние плода было очень тяжелым, сердцебиение отсутствовало. Проводились массаж сердца и вентиляция легких,затем он был переведен в отделение интенсивной терапии, но на следующий день умер.
Раздел 2. Проблема оказания помощи
Мониторинг плода на первой и второй стадиях родов.
Клиническая политика и факторы больного
Болезненный и относительно короткий первый период родов. Сердцебиение плода трудно мониторировать. Головной электрод подключен в 7:15. Шейка раскрылась полностью в 8:05. Больная очень напряжена и не способна к совместным действиям. Рекомендована, но не выполнена эпизиотомия. Присутствие мужа на несколько минут. Выполнена эпизиотомия. Головной электрод удален после прорезывания головки в 8:15. Перед удалением электрод показал значительное урежение ритма сердцебиения плода.
Способствующие факторы
|
Специфические |
Общие |
|
Условия работы |
Нет |
|
Роддом находится в состоянии интенсивного ремонта, но продолжает функционировать. Нарушение нормальной географии | |
|
Факторы команды | |
|
Заметки не были найдены в библиотеке. План помощи, составленный консультантом, не был прочитан. Отделение укомплектовано персоналом, рабочая нагрузка средняя. |
Изменение процедур по дежурству, необходимость обеспечения быстрого восстановления записей. |
|
Индивидуальные факторы |
|
|
Акушерки не обратили внимания на замедление ритма сердца плода, потому что они были отвлечены. |
Тренинг использования головного электрода. |
|
Факторы задачи |
|
|
Акушерки не осознавали возможность дистоции. Задержка между прорезыванием головки и завершением родов. Головной электрод не защищен протоколом. |
Отсутствие ясного протокола действий. |
Факторы политики и управления учреждения
Отделение не имело заведующего акушерской службой 2 года.
Каждая из этих проблем оказания помощи была проанализирована в отдельности. Здесь указан только неадекватный мониторинг плода. Многие способствующие факторы повлияли на оказание помощи на этой стадии родов. Персонал столкнулся с больной, которая находилась в крайне напряженном состоянии, не принимая их рекомендации. Головной электрод был удален без согласования с отделенческим протоколом, акушерки были растеряны из-за состояния матери, план помощи, составленный консультантом, не был рассмотрен, работа родильного отделения нарушена из-за ремонтных работ. Лишь некоторые из этих факторов оказали общее влияние на работу отделения, особенно доступность протокола, обучение кардиотокографии и порядка удаления головного электрода.
Принципы риска и безопасности
Приведенный выше анализ человеческой ошибки, модели и рамок организационного происшествия охватывает многие стороны контроля риска и улучшения безопасности.
Персональный и системный взгляды на безопасность
Существует давняя и широко распространенная традиция личностного подхода к расследованию небезопасных действий, погрешностей и грубых нарушений в выполнении процедур людьми переднего края работы (медсестрами, терапевтами, хирургами, анестезиологами и т.п.) (13). Считается, что эти небезопасные действия вызваны такими индивидуальными факторами, как забывчивость, невнимательность, слабая мотивация, беззаботность, неряшливость, безрассудность. Лечение направлено, главным образом, на индивидуальное поведение. Эти методы включают написание новых процедур (или дополнение существующих), дисциплинарные меры, прекращение споров, переобучение, название имен, обвинения и пристыжение. Последователи этого метода предпочитают относиться к ошибкам как к моральным проблемам, считая, что плохие вещи случаются с плохими людьми.
Базовые предпосылки системного подхода, описанного здесь, - это то, что людям свойственно ошибаться, даже лучшие люди, работающие в лучших организациях, допускают погрешности. Погрешности оказываются, скорее всего, следствием, а не причиной, имеющим в своей природе больше системных «восходящих» факторов, а не порочность человеческой натуры. Они включают повторяющиеся «западни» погрешностей рабочего места и организационного процесса. Меры противодействия основываются на том, что мы не можем изменить состояние человека, но можем изменить условия работы человека (13). Меры, направленные на индивидуальное (обучение), остаются важными, но недостаточными.
Расследование и анализ
В здравоохранении развивается этот метод расследования. Хотя для серьезного следствия необходимы клиническая интуиция и опыт, многое достигается систематическим подходом и большим вниманием к организационной политике, чтобы выявить проблемы оказания помощи. В выявлении способствующих факторов и выяснении, что каждая проблема оказания помощи может иметь различный набор способствующих факторов, - задачи незнакомые. Процесс может показаться сложным, требующим времени, однако, мы видим, что использование этой системы ускоряет сложные расследования путем фокусирования на ключевых темах, избежание рутинной раздачи обвинений и выделение факторов, на которые нацелено расследование.
Вмешательства, улучшающие безопасность
Как может выглядеть программа безопасности в здравоохранении? В промышленности программа безопасности нацелена скорее на задачи, команду, условия работы, чем на персонал (14). Безопасность должна адресоваться и на частный клинический процесс (клинический аудит), и на межличностный и организационный уровни. Для аудита необходим широкий анализ организационных и системных особенностей. Когда задача ясно определена, тогда возможна большая стандартизация, ясный протокол действия, меньшее доверие к капризам человеческой памяти. Командные и коммуникационные нарушения обязательно рассматриваются при анализе большинства происшествий, а меры их решения могут быть простыми и узконаправленными. Системы, применяемые в промышленности, мониторируют условия работы и связанные с ними организационные факторы и вызывающие эти условия решения. Условия, предрасполагающие к риску и небезопасной практике, должны находится под рутинным наблюдением, оценивая не здоровье больного, а здоровье отделения, т.к. эти факторы являются его признаками жизни.
Изучение успеха
Хотя мы и выделяем большую проблему человеческой ошибки и риска больного, не все решения приходят из изучения плохой работы. Например, натуралистические исследования и личный клинический опыт показали значительные способности людей отвечать на стресс, быстро и эффективно использовать экспертные знания, быстро адаптироваться к изменяющимся условиям. В хирургии исследования направлены и на человеческие ошибки, и на адаптацию людей. Например, de Leval et al (15) в исследовании операций на артериях выявили множество этапов, потенциально угрожающих жизни. Однако, результаты говорят, что на исход оказывало влияние не возникновение проблем, а способность хирурга их компенсировать. Хирургическое совершенство достигается не совершением ошибок, но пониманием того, что ошибки могут допускаться и необходимо развивать методы их компенсации.
От контроля риска к открытой культуре безопасности
В будущем возникнет множество возможностей для контроля риска и безопасности больного. Один фактор может оказать длительное влияние на контроль риска, ответственность за который несет небольшое число людей в каждом учреждении здравоохранения. В авиации, наоборот, «безопасность - ответственность каждого». В медицине каждый заботится о безопасности больного, желая сделать это лучше. Однако, необходимо ввести безопасность больного в культуру здравоохранения, не только как чувство индивидуального высокого стандарта, но и широкое принятие необходимости открытого признания ошибок и отрицательных явлений, системное понимание риска и безопасности, необходимость для каждого активно способствовать безопасности больных.
Литература
1. Kohn LT, Corrigan JM, Donaldson MS, eds. To err is human: building a safer health system. Washington DC: National Academy Press, 1999.
2. Wilson KM, Runciman WB, Gibberd RW, Harrison ВТ, Newby L, Hamilton JD. The Quality in Australian Health Care Study. Medical Journal of Australia 1995; 163:458-471.
3. Department of Health. An organisation with a memory. London: HMSO, 2000.
4. Leape L, Berwick D. Safe healthcare: are we up to it? British Medical Journal 2000; 320:725-6.
5. Chantler С The role and education of doctors in the delivery of health care. Lancet 1999; 353:1178-81.
6. Reason JT. Human error. New York: Cambridge University Press, 1990.
7. Reason JT. Adverse events: the human factor. In: Vincent CA, ed. Clinical risk management. Enhancing patient safety. London: British Medical Journal Publications, 2001.
8. Vincent CA, Taylor-Adams S, Stanhope N. Framework for analysing risk and safety in clinical medicine. British Medical Journal 1998; 316:1154-1157.
9. Stanhope N, Vincent CA, Taylor-Adams S, O'Connor A, Beard R. Applying human factors methods to clinical risk management in obstetrics. British Journal of Obstetrics and Gynaecology 1997; 104:1225-32.
10. Taylor-Adams SE, Vincent C, Stanhope N. Applying human factors methods to the investigation and analysis of clinical adverse events. Safety Science 1999; 31:143-59.
11. Vincent CA, Taylor-Adams S, Chapman EJ, Hewett D, Prior S, Strange P, Tizzard A. How to investigate and analyse clinical incidents: Clinical Risk Unit and Association of Litigation and Risk Management protocol. British Medical Journal 2000; 320: 777-81.
12. Vincent CA, Taylor-Adams S, Chapman EJ, Hewett D, Prior S, Strange P, Tizzard A. The investigation and analysis of clinical incidents: a protocol. London: Royal Society of Medicine Press, 1999.
13. Reason JT. Human error: models and management. .British Medical Journal 2000; 320:768-70.
14. Vincent CA. Risk, safety and the dark side of quality. British Medical Journal 1997; 314:1775-1776.
15. De Leval, MR, Carthey J, Wright DJ, Farewell VT, Reason JT. Human factors and cardiac surgery: a multicentre study. Journal of Thoracic Cardiovascular Surgery 2000; 119:661-672.
16. Vincent CA, ed. Clinical risk management. Enhancing patient safety. London: British Medical Journal Publications 2001. To be published November 2000 (www.clinicalrisk.bmjbooks.com)
АНЕСТЕЗИОЛОГАМ ОБ ОКСИДАЗНОМ СТРЕССЕ, АПОПТОЗЕ И НЕЙРОТРАНСМИССИИ
М.Лэйми, Д. Деби Дюпон (Льеж, Бельгия)
Введение
Нейротрансмиссия и апоптоз, а также роль оксидазного стресса становятся яснее, но много вопросов остаются не понятными до конца, и прежде всего, «оксидазный стресс». Мы представляем обзор современных достижений в области нейротрансмиссии и механизмов апоптоза, а также роли активных радикалов кислорода (АРК).
Основные механизмы нейротрансмиссии и апоптоза
Передача сигнала информации зависит от наличия испускающей сигнал клетки и воспринимающей клетки (рис.1). Передача информации может осуществляться в виде электрического сигнала, химических молекул, межклеточных соединений в виде контактных ворот, взаимодействий поверхностных лигандов и рецепторов и т.д. (1).
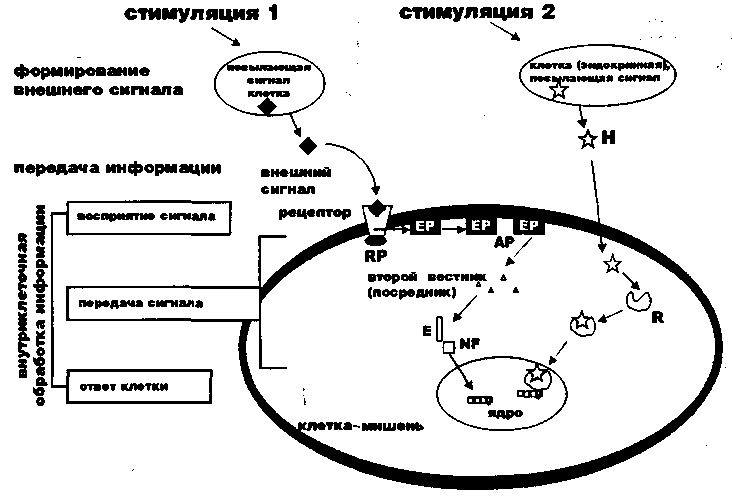
Рисунок 1. Схема передачи информации от внешних источников сигнала до клетки мишени. Есть два основных пути получения информации от внешних источников: посредством рецептора (стимуляция 1) и сигнал Н, проходящий в клетку путем диффузии (стимуляция 2) (детализация -в тексте). RP - цитоплазматический или ядерный рецептор; ЕР - эффекторные белки; АР - воспринимающие белки; Е - ферментный каскад; NF - ядерные факторы.
Клеточная обработка стимулов включает в себя синтез внешнего сигнала, перенос этой информации до клетки-мишени, его непосредственную обработку этой клеткой путем биотрансформации химических реакций на протяжении от рецептора до завершения клеточной реакции на полученный сигнал. Все эти этапы тщательно контролируются, а клетка, испускающая сигнал, четко зависима от регулирующих ее состояние метаболизма, повторения информации, ее диференцировки и апоптоза воспринимающей клетки.
Описано два варианта входа внешнего сигнала в воспринимающую клетку: пассивная диффузия и рецепторное связывание. При пассивной диффузии посредник (Н - чаще всего гормоны) связывается с цитоплазматическими или ядерными рецепторами (рис.1, путь стимуляции 2). Если внеклеточный посредник не диффундирует в клетку-мишень, то происходит его взаимодействие с рецепторами наружной мембраны клетки (рис.1, путь стимуляции 1), в том числе и модифицирующими рецепторами (чаще - путем олигомеризации). Затем сигнал переносится внутрь клетки при участии трансмембранной части рецептора и/или белков, сцепленных с цитоплазматической частью рецептора (ЧР), и работающих по типу «включателя» при дальнейшей передаче сигнала. Внутри клетки наступает каскадный механизм обработки информации, проходящей через эффекторные (ЕР) и воспринимающие белки (АР), второго посредника и ферментативный каскад (Е). При этом в цитоплазматическом комплексе вырабатываются ядерные факторы (NF) и ингибиторы вслед за транслокацией ЯФ в ядро, где они связываются с соответствующим участком ДНК, запуская процесс транскрипции специфических генов. Финальный результат проявляется в виде активации клеточных белков, запускающих многообразные реакции: иммунные, воспалительные, а также реакцию программированной гибели клетки, известную как «апоптоз» (1,2). Очень много неясности в механизмах механической ориентации клетки, выражающейся в избирательной активации генов или апоптозе, но благодаря последним сведениям стало известно, что участие некоторых ядерных факторов более специфично при апоптозе, чем других.
Внутриклеточные компоненты каскадной обработки сигнала
Компонентами внутримолекулярной обработки и передачи сигнала являются белки или небольшие молекулы, часто называемые посредниками второго вестника (циклические нуклеотиды, трифосфат инозитола и ионы Са++), которые активируются и запускают соответствующие ферменты передачи сигнала. Белки, которые участвуют в таких процессах, обычно являются гормонами, эффекторные белки (чаще - связанные с клеточной мембраной) являются запускающими в процессе передачи сигнала и АР, которые в основном несут связующую роль между исключительными компонентами передачи сигнала. Эти АР являются также и местом гибели или нейтрализации многих переносящих информацию элементов.
Краеугольным камнем регуляции процесса передачи информации является процесс фосфорилирования и протеолиза. Фосфорилирование осуществляется на се-рин/треониновом или тирозиновом окончании за счет протеинкиназ, что создает возможность появления новых точек связывания белка. Фосфорилирование белков регулируется дефосфориляцией белковыми фосфатазами (2,3).
Протеолиз вмешивается в процессы зарождения новых белков (активация проферментов путем ограниченного протеолиза) и их деградации. Образование протеасом – основной путь протеолиза в процессе передачи сигнала. Протеасомы представляют собой крупные многобелковые комплексы, которые разрушаются путем вездесущего фосфорилирования. Роль протеасом хорошо описана в реакциях деградации комплексов, сформированных за счет NF и их ингибиторов (4).
Ядерные факторы
Ядерные факторы присутствуют в цитоплазме в виде неактивных форм, сцепленных со специфическими ингибиторами. Фосфорилирование - основной механизм, контролирующий их активность. Их выброс мониторируется путем оксиметрии в луковице яремной вены; оксигенация ткани головного мозга (PO2) ингибирует и регулирует их транспорт в ядро клетки. После димеризации они связываются со специфическими элементами ДНК и действуют как активаторы или репрессоры процесса считывания информации в комплексе с другими регуляторными белками. Семейство ядерных факторов состоит из ЯФ-кВ/Rel факторов транскрипции и представляет собой пять различных, связанных с элементами ДНК, субъединиц: ЯФ-кВ 1/Rel (р50), ЯФ-кВ2/Реl (р100/р52), Rel A (p65), Pel В и c-Rel (5). У родственных образований присутствует гомологичный домен из ± 300 аминокислот, ответственных за связывание с IkB протеином, димеризацию и связывание с аминокислотами. Они также сплетаются в гомологичные или гетерологичные димеры, сосредоточенные в цитозоле в виде неактивных форм и связанных с молекулами-ингибиторами семейства IkB. Одним из представителей этого ряда является гетерологичный димер р50:р65 (Rel А), обозначаемый как ЯФ-kB. Это уникальный фактор в силу его быстрой активации и регуляторной способности. Он играет главную роль в воспалительном и иммунном ответе, когда необходима быстрая активация гена для сохранения жизни клетки (продукция цитокинов и протеинов острой фазы). В большинстве типов клеток этот механизм препятствует апоптозу. ЯФ-кВ активируется основными стимуляторами (провоспалительными цитокинами, бактериями, ЛПСД, вирусами), а также реактивными продуктами распада кислорода, которые образуются в ходе воспалительной реакции. Его активация начинается с процесса фосфорилирования на двух сериновых окончаниях IkB, активируемого специальной lkB-киназой (IKK). Затем реакция распространяется на всю субстанцию, приводя к быстрой деградации IkB протеасомами (4). ЯФ-кВ вырабатывается и быстро транслоцируется в ядро. Регулирование активации ЯФ-кВ контролируется в основном интенсивностью и распространенностью реакции фосфорилирования 1кВ, а также и степенью активности протеасом (рис.2).

Рисунок 2. Активация ядерного фактора ЯФ-кВ в цитоплазме и внутриядерная активность. р50 и р65 (Rel А): субъединицы ЯФ-кВ; IkB : ингибитор ЯФ-кВ; IKK: специализированная протеин киназа, которая связывает 2 фосфата (Р) на двух сериновых окончаниях IkB. Фосфорилирование сменяется распространением реакции и диссоциацией ЯФ-кВ-lkB комплекса, которая позволяет ЯФ-кВ войти в ядро и запустить процесс транскрипции ДНК.
Специфические аспекты апоптоза
Апоптоз (программируемая гибель клетки: ПГК; часто называется как самоубийство клетки) запускается естественными и патологическим факторами и характеризуется запрограммированной относительно медленной морфологической эволюцией клетки, для которой необходим синтез новых матричных РНК и новых протеинов (2,6). Апоптоз это необходимый процесс для развития и обновления тканей, поддержания иммунного гомеостаза за счет обновления популяций лимфоцитов и элиминации определенных для этого и поврежденных клеток или избыточного количества лимфоцитов после удаления антигенов.
Апоптоз коренным образом отличается от некроза, который представляет собой острый процесс гибели клетки путем быстрого разрыва клеточной мембраны и дезинтеграции всей клетки. Апоптоз запускается состоянием ядерного материала, сжатием и обезвоживанием клетки, которые заканчиваются ее фрагментацией и выбросом пузырьков, апоптических тел. Последние захватываются фагоцитами без сопровождения воспалительной реакции (рис.3). Недостаточность или избыток апоптоза играет значительную роль в патогенезе таких заболеваний, как рак или аутоиммунные болезни, а также участвует при функциональных нарушениях, сопряженных с процессами ишемии-реперфузии (7). Апоптоз запускается специфическими сигналами и часто зависим от их передачи, но не всегда, так как в клетке уже присутствуют крупные части сигнальной субстанции в латентной форме, которые могут быть активированы простой стимуляцией.
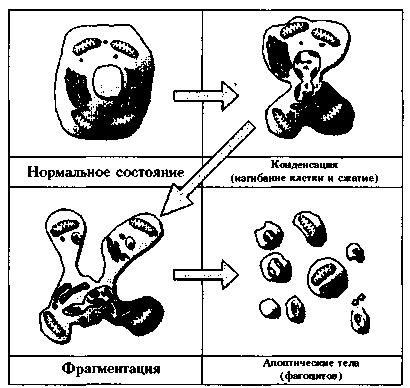
Рисунок 3. Схематическое изображение процесса апоптоза клетки: конденсация ядерного вещества, сжатие и формирование изгибов клетки, фрагментация и выброс пузырьков (апоптических тел).
Ключевыми элементами апоптоза являются гибель имеющихся белков посредством протеолиза и реакция расщепления caspases (цистеин-аспартат протеаза, ЦАП), используемая в этом протеолизе (8,9,10). Эти ферменты (14 разных энзимов млекопитающих) присутствуют в неактивных формах (зимогены) и активируются (самостоятельно при участии ЦАП) с образованием энзимов-тетрамеров. Две группы ЦАП отличаются по структуре зимогенов: ЦАП инициатор и ЦАП эффектор, причем последний активируется предыдущим. Есть сообщения о существовании трех подгрупп ЦАП, активируемых исключительно при воспалении (ЦАП 1,4 и 5: подгруппа 1) или при апоптозе (ЦАП 3,6,7,8,9 и 10: подгруппы 2 и 3), однако такая классификация остается пока в стадии обсуждения. При апоптозе участвуют 2 каскада ЦАП: каскад, запускаемый при гибели рецепторов на поверхности мембраны, и каскад, инициируемый стрессовой альтерацией в митохондриях. Первый каскад инициируется при связывании погибшего сигнального участка с рецептором гибели на поверхности мембраны клетки (рис.4).

Рисунок 4. Схематическое изображение каскада апоптоза, начинающегося с активации мембранного рецептора: цепочка запуска рецептора гибели клетки и активация «эшафота» ЦАП, Связанный с лигандом (Fas-L) и собственно гримерный рецептор Fas (CD95) взяты в качестве типичного примера трансмембранного рецептора. Он связывается посредством домена гибели клетки (DD) с ассоциируемым протеином (FADD), который представляет собой эффекторный домен гибели клетки (DED), и вовлекает в процесс инициатор проЦАП.
Такая цепочка связывания индуцирует олигомеризацию рецепторов, задействует внутриклеточные белки, которые связаны с рецепторами, формирующими комплекс, содержащий информацию (сигнал) о гибели клетки (DISC) и запускающий «эшафотную активацию» каскадной цепочки ЦАП, начиная с ЦАП-8. Вторая каскадная цепочка начинается со стимуляции (обычно присутствующей при стрессе), которая не приводит к связыванию рецепторов, но заставляет митохондрии выбрасывать цитохромы (рис.5).
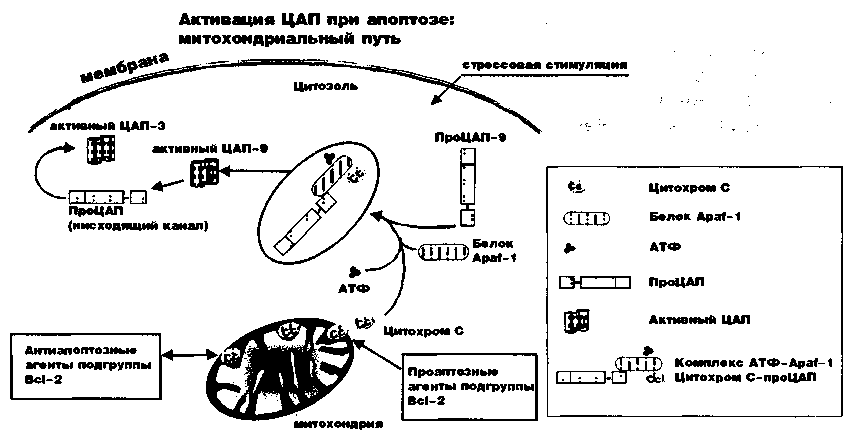
Рисунок 5. Митохондриальный путь апоптоза: стрессовая стимуляция нарушает триггерный I выброс митохондриями цитохрома С (Cyt с). С этого момента апоптоз развивается механически. Цитохром С формирует комплекс со специальным белком Apaf-1, связываясь при этом с АТФ и вовлекая проЦАП-9.
Проапоптозные и антиапоптозные агенты подгруппы Bcl-2 регулируют выработку цитохрома С. В цитоплазме цитохром С формирует комплекс с цитоплазматическими фаЙ-торами и специальным белком Apaf-1, который связывается и гидролизирует АТФ. Цитохром C-Apaf-1 комплекс вовлекает, связывает и активирует ЦАП-9 (восходящий каскад), который в последующем активирует ЦАП-3,6 и 7 (нисходящий каскад).
На различных уровнях каскадной цепочки апоптоза действуют различные механизмы регуляции, но процессами регуляции активации каскада являются фосфорилирование и дефосфорилирование протеинкиназами и фосфатазами.
Роль реактивных радикалов кислорода в передаче сигнальной информации при апоптозе.
Реактивные радикалы, образующиеся из кислорода (РРК) и из соединения азота с кислородом (РРАК), играют критическую роль в механизме воспаления при септицемии и респираторном дистресс синдроме. РРАК являются мощными противомикробными агентами, заставляющими микроорганизмы разрушаться до фаголизосом, но они также являются и повреждающими агентами близлежащих клеток и тканей, так как вырабатываются бесконтрольно стимулируемыми фагоцитами на фоне острой воспалительной реакции. За их продукцию и разрушение ответственны специфические ферменты, которые могут быть идентифицированы и характеризованы (рис.6)

Рисунок 6. РРАК каскад, начинающийся с супероксид аниона fO^'J и оксида азота (NO), запускается соответствующими ферментами. Образуется пероксинитрит (ONOO-), нестабильное вещество, реагирующее со многими биомолекулами (особенно с -SH группами (NO2~ - нитрил-радикал и ОН-гидроксил-радикал) или с нитрониум ионом (NO2+), ответственными за процессы нитрирования и гидроксилирования. Перекись водорода (H^Gg) образуется из радикала кислорода (U2~ ); при наличии комплексиона железа (+L-Fe2+ ) из перекиси водорода образуется гидро-ксильный радикал (-ОН), а в полиморфноядерных фагоцитах при участии миелопероксидазы (МРО), перекиси водорода и хлорид аниона (Ch ) образуется солянистая кислота (HOCI), которая ответственна за окисление клеточных структур. Она реагирует с нитритом (NO2~, который образуется в результате реакции радикала NO и молекулярного кислорода О2 ) и пероксинитритом (ONOO~), в результате чего образуется нестабильный нитрил хлорид (NO2CI), распадающийся на NO2 • -радикал и чистый хлорид).
РРАК могут присутствовать во многих типах клеток (лимфоциты, моноциты, макрофаги, нейтрофилы) и их образование является основным in vivo феноменом, который точно регулируется и обуславливает нормальный клеточный метаболизм. Основными эндогенными источниками РРАК являются продукты активации реакций с участием таких ферментов, как НАДФ-оксидаза, NO-синтетаза, ксантин-оксидаза, цепочка транспорта электронов в митохондрии, метаболизм арахидоновой кислоты и активность цитохрома Р450. В норме однако РРАК образуется немного. В 1973 году было определено, что в нормальных условиях митохондриями расходуется ±2% кислорода на построение супер-оксид иона (О2-) и его продукта дисмутации - перекиси водорода. Это наблюдение имело важное значение для продолжения исследований по передаче сигнальной информации в клетке (11).
Оксидазный стресс и окислительно-восстановительный баланс
Состояние компонентов окисления-восстановления (Е0) определяется возможностью захвата (окислительный компонент) или выброса (восстановительный компонент) одного или более электронов. Для нормальной биологической среды характерно наличие равновесия между оксидантами и восстановителями (РОВ). Несмотря на определенные трудности в точном измерении этого РОВ, цитоплазма в физиологическом состоянии представляется более основной средой за счет присутствия в ней глутатиона (ГТИ), НАД/НАДФ, особенно в лейкоцитах (рис.7).
Нарушение РОВ внутри клетки немедленно влечет за собой запуск механизмов переноса сигнальной информации и главная роль в этом принадлежит функции -SH групп. Функциональная активность тиоловых групп зависит от множества компонентов, участвующих в процессе переноса сигнальной информации: клеточных рецепторов, ЦАП, ядерных факторов и даже ядерных белков, которые регулируют связывание ЯФ с элементами ДНК. Функции -SH групп легко нарушаются в результате окислительных реакций и являются идеальной мишенью РРАК. Физиологическая роль РРК заключается в изменении РОВ за счет потребления основных молекул (ГТИ, НАД, НАДФ) или в результате оксидазной модификации структуры белков. Окислительное повреждение белков позволяет последним взаимодействовать с рецепторами, изменять истинную ферментную активность или связываться со специфическими мишенями на ДНК, запуская, таким образом, перенос сигнальной информации и генные проявления. В лейкоцитах нарушения РОВ можно представить изменением переноса сигнальной информации.
В физиологических условиях баланс ферментных и неферментных антиоксидантов создает РОВ, задерживая окислительные и восстановительные изменения внутри клетки. Этот баланс нарушается избыточной активацией НАДФ-оксидазы в стимулированных лейкоцитах. In vivo избыточная продукция реактивных радикалов кислорода и азота возможна при воспалении, для которого характерна интенсивная активация лейкоцитов, или при таких необычных ситуациях, как массивная радиация. В таких случаях, когда буферная емкость антиоксидантов становится избыточной или патологически снижается, возможны оксидазные повреждения с развитием оксидазного стресса.