

Функции глутамата в нервной ткани
- Энергетическая: глутаминовая кислота связана большим числом реакций с промежуточными метаболитами цикла трикарбоновых кислот;
- участие (вместе с аспартатом) в реакциях дезаминирования других аминокислот и временном обезвреживании аммиака;
- образование из глутамата нейромедиатора ГАМК;
- участие в синтезе глутатиона — одного из компонентов антиоксидантной системы организма.
Глутамат с большой скоростью образуется из своего кетоаналога — -кетоглутаровой кислоты в ходе реакции трансаминирования и используется для образования глутатиона, глутамина и -аминомасляной кислоты.
Ткань мозга способна синтезировать заменимые аминокислоты.
Образование аммиака происходит в пуриновом цикле: путем гидролитического дезаминирования АМФ образуется ИМФ и аммиак. ИМФ, далее конденсируясь с аспарагиновой кислотой, образует аденилсукцинат, который, расщепляясь, образует вновь АМФ (и фумарат). Фумарат в ЦТК образует ОАА, который поддерживает уровень аспарагиновой кислоты, вступая в реакцию переаминирования с глутаминовой кислотой.
Биохимические основы возникновения и проведения нервного импульса
В норме в состоянии покоя мембрана аксона поляризована: внутри аксона ионов калия в 30 раз больше, чем ионов натрия. Концентрация анионов также различна. Катионы внутри клетки нейтрализуются в основном белками и фосфатами, которые не могут выходить наружу; внеклеточные катионы (в основном Na+) уравновешиваются Cl–, проницаемость которого выше, чем белков. Это во-первых. Во-вторых проницаемость Na+ составляет 1/20 проницаемости К+. В третьих, К+, Na+ АТФ-аза выкачивает 3 Na+ в обмен на 2 К+. При таких условиях внутренняя сторона клеточной мембраны заряжена электроотрицательно по отношению к наружной поверхности и электродвижущий трансмембранный потенциал Е=-70 мВ. При действии медиатора происходит активация аденилатциклазы мембран, под влиянием которой из АТФ образуется ц-АМФ, включающий каскадный механизм активации (фосфорилированием белков) натрий-калиевой АТФ-азы (ионного насоса). Таким образом, с участием АТФ происходит вначале открытие каналов для прохождения ионов натрия внутрь аксона, а затем открытие каналов для выхода ионов калия наружу. В этом состоянии разность потенциалов достигает +40мВ, положительный заряд внутри аксона. Это потенциал действия и, возникнув в одном участке, вследствие диффузии ионов вдоль волокна, снижает потенциал покоя и вызывает здесь тоже развитие потенциала действия. Возникает волна деполяризации — нервный импульс; затем происходит распространение волны деполяризации. Восстановление мембраны в поляризованном виде происходит также с участием ионных насосов с затратой АТФ. Следовательно, для функционирования нервной системы необходима выработка и затрата значительных количеств АТФ.
Понятие о синапсах. Нейромедиаторы. Пептиды
Синапс - функциональный контакт специализированных участков плазматических мембран двух возбудимых клеток. Состоит из пресинаптической мембраны, синаптической щели и постсинаптической мембраны. Мембраны клеток в месте контакта имеют утолщения в виде бляшек — нервных окончаний. Нервный импульс, достигший нервного окончания, не в состоянии преодолеть возникшее перед ним препятствие — синаптическую щель. Здесь электрический сигнал преобразуется в химический. Пресинаптическая мембрана содержит специальные канальные белки, которые реагируют на мембранный потенциал, изменяя свою конформацию, и формируют канал. В результате ионы Са2+ проходят через пресинаптическую мембрану по градиенту концентраций в нервное окончание. Градиент концентраций Са2+ создается работой Са2+ -зависимой АТФазы — кальциевым насосом. Повышение концентрации Са2+ внутри нервного окончания вызывает слияние 200-300 имеющихся там везикул, заполненных ацетилхолином, с плазматической мембраной. Далее ацетилхолин секретируется в синаптическую щель путем экзоцитоза и присоединяется к рецепторным белкам, расположенным на поверхности постсинаптической мембраны.
При взаимодействии с ацетилхолином белок-рецептор изменяет свою конформацию, формируя натриевый канал. Катионная селективность канала обеспечивается тем, что ворота канала сформированы отрицательно заряженными аминокислотами. Таким образом, повышается проницаемость постсинаптической мембраны для натрия и возникает новый импульс (или сокращение мышечного волокна). Деполяризация постсинаптической мембраны вызывает диссоциацию комплекса «ацетилхолин - белок - рецептор», и ацетилхолин освобождается в синаптическую щель. Как только ацетилхолин оказывается в синаптической щели, он подвергается быстрому гидролизу под действием ацетилхолинэстеразы. Образуется промежуточный фермент-субстратный комплекс, в котором ацетилхолин связан с активным центром фермента через серин.
Нейромедиаторы - вещества, характеризующиеся следующими признаками:
- накапливаются в пресинаптической структуре в достаточной концентрации;
- освобождаются при передаче импульса;
-вызывают после связывания с постсинаптической мембраной изменение скорости метаболических процессов и возникновение электрического импульса;
-имеют систему для инактивации или транспортную систему для удаления из синапса, обладающие к ним высоким сродством.
Нейромедиаторы обеспечивают синаптическую передачу нервного импульса. Их синтез происходит в теле нейронов, а накопление — в особых везикулах, которые постепенно перемещаются с участием систем нейрофиламентов и нейротрубочек к кончикам аксонов.
К нейромедиаторам относят аминокислоты (и их производные): таурин, норадреналин, дофамин, ГАМК, глицин, ацетилхолин, гомоцистеин, адреналин, серотонин, гистамин.
Таурин. Образуется из аминокислоты цистеина. Сначала происходит окисление серы в SН-группе до остатка серной кислоты (процесс идет в несколько стадий), затем декарбоксилирование. Таурин не имеет карбоксильной группы, но содержит остаток серной кислоты. Участвует в проведении нервного импульса в процессе зрительного восприятия.
Ацетилхолин. Синтезируется из холина и Ацетил-КоА. Участвует в синаптической передаче нервного импульса. Накапливаясь в синаптических пузырьках, образует комплексы с отрицательно заряженным белком везикулином. Передача возбуждения с одной клетки на другую осуществляется с помощью специального синаптического механизма.
Катехоламины: норадреналин и дофамин. Синтезируются по общему механизму из тирозина. Ключевой фермент синтеза — тирозингидроксилаза, ингибируемая конечными продуктами. Норадреналин — медиатор в постганглионарных волокнах симпатической и в различных отделах ЦНС.
Биосинтез катехоламинов
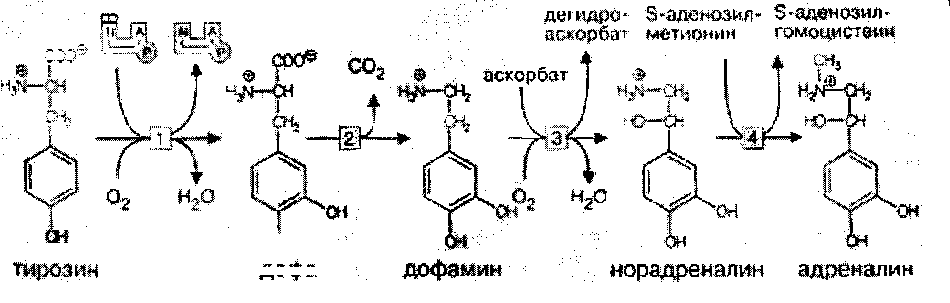
Дофамин — медиатор проводящих путей, тела нейронов которого расположены в отделе мозга, отвечающем за контроль произвольных движений.
Катехоламины, как и ацетилхолин, накапливаются в синаптических пузырьках и выделяются в синаптическую щель при поступлении нервного импульса. В пресинаптической мембране имеется специальный регуляторный белок —-ахромогранин, который в ответ на повышение концентрации медиатора в синаптической щели связывает уже выделившийся медиатор и прекращает его дальнейший экзоцитоз. Фермента, разрушающего медиатор, в адренэргических синапсах нет. После передачи импульса молекулы медиатора перекачиваются специальной транспортной системой путем активного транспорта с участием АТФ обратно через пресинаптическую мембрану и включаются вновь в везикулы. В пресинаптическом нервном окончании излишек медиатора может быть инактивирован моноаминоксидазой или катехоламин-О-метилтрансферазой путем метилирования по оксигруппе. Передача сигнала в адренэргических синапсах происходит с участием аденилатциклазной системы. Связывание медиатора с постсинаптическим рецептором почти мгновенно вызывает повышение концентрации ц-АМФ, что приводит к быстрому фосфорилированию белков постсинаптической мембраны. В результате тормозится генерация нервных импульсов постсинаптической мембраной. МАО (моноаминоксидазы).
ГАМК — тормозной медиатор. Повышает проницаемость постсинаптических мембран для ионов калия, вызывая изменение мембранного потенциала.
Глицин — тормозной медиатор, по вызываемым эффектам подобен ГАМК.

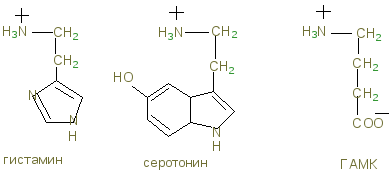
Нейромедиаторы и нейрогормоны
